�����ĺ�Ĭ��(Alzheimer��s disease, AD)��������Ϊ���书�ܵ�ɥʧ�����ұ���Ϊ˯�������������������������֪�ϰ�(mild cognitive impairment, MCI)�ξ��Ѿ������ԡ�˯��ʱƤ������(slow oscillations, SO��0.5-1 Hz)������Ƥ��Ĵ���(12-15 Hz)�Լ����ǵ�ʱ��Э���Ա���Ϊ�Ǽ����γɵĹؼ��������о�������ֱ����̼�(slow oscillatory transcranial direct current stimulation, so-tDCS)��DZ�����ô̼���˯��״̬�����ķ�ʽ�ڰ���С˯�ڼ�Ӧ�ã��Ե���9�����Ժ�7��Ů��MCI���ߵ���Щ�ģʽ����˯����صļ��乮�̡��̼�������������SO(����)�ͷĴ����ʣ���SO�����ηŴ��˷Ĵ����ʣ�������EEG��¼��SO�ͷĴ����ʲ���֮���ǿ��ͬ���ԡ����⣬��ٴ̼���ȣ�so-tDCS�������Ӿ������Լ��䣬�����Ӿ������Լ������ǿ��ͬ������ء���Щ����ΪMCI���ߵ�˯�������ϰ��ͼ���ȱ���ṩ��һ�����������õ����Ʒ��������ٽ������Ƕ������乮�̵����⡣���ķ�����The Journal of Neuroscience��־��(�������ź�siyingyxf��18983979082��ȡԭ�ģ���˼Ӱ�ṩ����������ط�������ҪҲ�����Ӵ��ź���Ⱥ��ԭ��Ҳ����Ⱥ���)��
�¹ʶ�֪�£�������������ؽ���Ķ���ֱ�ӵ����������������ź�siyingyxf��18983979082��ȡԭ�ļ������������
ͨ�����ļд������������
���Ƴ�40Hz tACS�����ĺ�Ĭ�����ߺ�����ע��Ӱ��
ͨ���Ե�ͼ/�Դ�ͼ�۲쵽�Ĵ��Ի��ָ�����Դ̼�
TMS-EEG���ٴ�Ӧ�ü�չ��
�����ĺ�Ĭ���������Դ̼�
������Ϣ�������Դ̼�����
�����������ϵ����Ӻͼ��ٻ���Ӧ���Ӻͼ�����������е��
����Ƶ���������֪���Ʋ�ͬ�ɷ��е��������
˯�ߡ����Դ̼����ϻ��Ĵ����о�
Nature Medicine����������̼����Ը���ǿ��֢
����̼��������Ͳ���˥�Ϲ������龰�����Ӱ��
����TMS��EEG��MRI���������������
ʹ�ô̼��豸����·���ز���Ծ�����������
�Ⱥͦ����ڹ���������������õ����֤��
TMS-EEG�о������Է�ӦΪ���к���˶��ָ��ṩ���廯����
Ƥ���˶��˷��Բ���������mu������λ�ĵ���
TMS�CEEG���Ϸ������������Ƥ��������̽���еĹ���
ʵʱEEG������TMS������֢���������ǰ��ҶƤ���������ͬ���̼�
��������̼���tACS�������������˹�������Ļָ�
tACS���EEG�о�:������������
��������̼�(tACS)��ʹ���Խ���ͬ���������֪����
1. ����
�ڰ����ĺ�Ĭ��(Alzheimer��s disease, AD)���ڣ��γɺͻָ��龰����������ѣ����ҹ����˸ü����ĺ�����ɲ��֡�ͬ����˯���ϰ��ƺ���AD��سմ���һ����Ҫ�������������������֪�ϰ�(mild cognitive impairment, MCI)���Ѿ��б�������MCIͨ����AD�մ������ס���Ȼ˯�������½�(��������乮����ص�˯�߲���)�ڽ����ϻ���Ҳ�ܳ���������MCI��AD�ͳմ����ߵ��½����س̶����Լӿ졣˯���жϲ����ᵼ��MCI���ߵļ������������DZ�ڲ�����չ����ֱ�����á�
˯���ڼ���ij��ڹ��������Ż������á�������˵������(slow oscillations, SO���������<1 Hz)��˯�߷Ĵ���(8-15 Hz)����ͨ��EEG�������ƺ��Գ����Լ���������Ҫ������������ϵͳ���̡����ۣ��±���ļ�����˯���ڼ䱻���¼�������ź������е������¼�(80-100Hz)����ͨ�����������Ƥ��֮���Э���Ի����·ֲ���Ƥ�㳤�ڴ洢���硣���ֶԻ�����Ƥ��SO������Ƥ����ٷĴ���(12-15 Hz)֮���������Ͻ鵼�ģ��Ĵ�����÷�����SO�����κͺ������ƾۼ��ڿ��ٷĴ����IJ��ȡ����Ĵ���(8-12 Hz)��˯�߷Ĵ������һ�֣����ڼ��乮���е����û���̫Ϊ����֪��
���˹��̷��棬Խ��Խ���֤�ݱ�����˯�ߣ��ر���SO�������������һ���ٽ�Ƥ�ʵ�������������������е�������������һ�ֲ���AD�������Ƶ��ġ���ˣ����˯�߲����ĸ�Ԥ�����ṩһ�����Ʒ������������ڼ��乮��ȱ�ݣ������ڽ��MCI���߰����ĺ�Ĭ���Ľ�չ������SO�����ϣ���İе��ѡ���˯���ڼ�Ӧ����������ֱ���̼�(slow oscillatory weak transcranial direct current stimulation, so-tDCS; Ƶ��< 1 Hz)�ṩ��һ����ǿSO��ķ������Է��������ڽ���������������������������������ִ̼�Ҳ�ᵼ�·Ĵ�������ӣ�����֪�����Ƿ�ı���SO�Ϳ��ٷĴ���֮��Ľ���Ƶ����ϣ��ⱻ��Ϊ��˯���ڼ���乮�̵Ĺؼ���so-tDCS���µļ�������ѱ�����֤������һЩ�о�û�з����κθ��ƻ������������෴��Ӱ�졣
����������¹ؼ����⣺
(1)˯�ߺͼ������������MCI�����Ƿ��Լ������˯���ڼ��so���Ĵ�����Լ����ǵĽ���Ƶ����Ϸ���������so-tDCS;
(2)so-tDCSЧӦ�Ƿ�����Ϊ�Ϸ�ӳ�ڼ�������С�Ϊ�˽����Щ���⣬������MCI������˯�ڼ�Ӧ����so-tDCS���������������Ӿ�ʶ��������EEG������˯�������ͼ�����ֵ�Ӱ�졣��ˣ����Ǽ�������ַ������Ը�Ԥ���Ƶ�DZ��������DZ�ڵĻ����ṩ���µļ��⡣
2. ���Ϻͷ���
2.1 ������
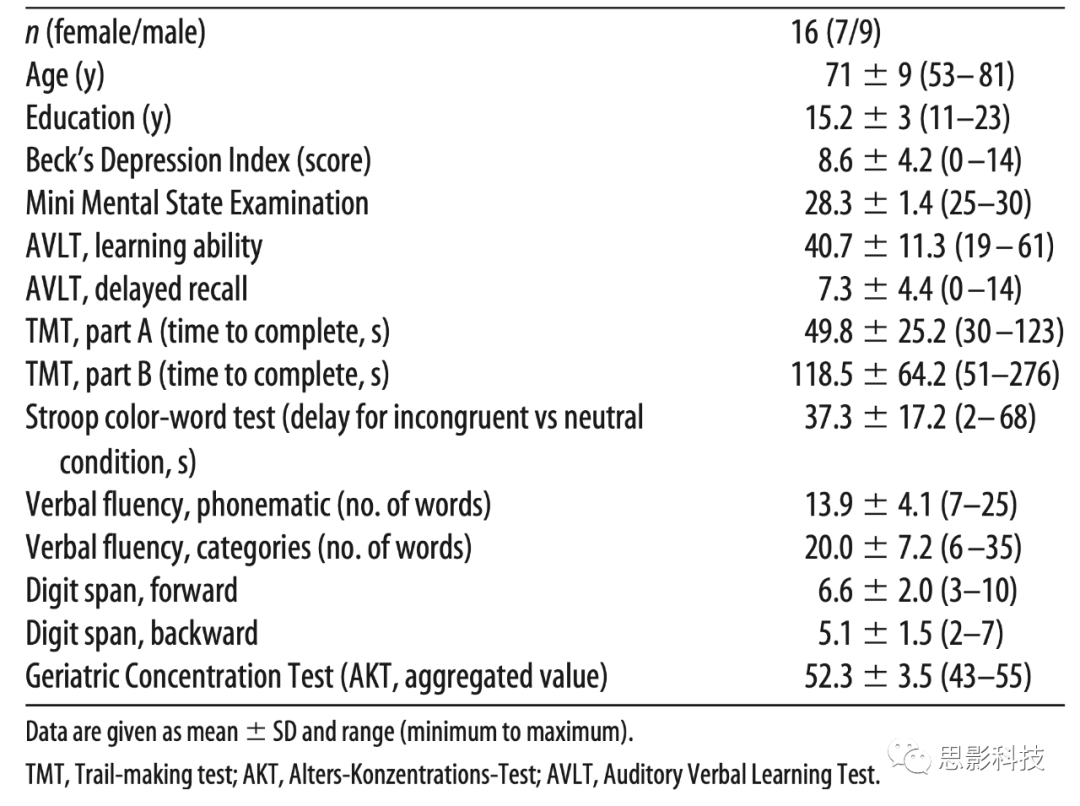
22��(10��Ů�ԣ�ƽ������71.2����8.79 SD����Χ��50-81��)�Ӳ����ش�ѧҽԺ���ڿƼ���������ļ��MCI���߲����˱��о���MCI����(�����ͣ�����Ͷ���)����Mayo Clinic�������ٴ���ϡ��÷����ٵ�����������̶�ƥ��Ľ����˵���������÷ֵ�1 SD (���������ں�����MCI)������Ӳ��ԣ��ܵ��ʱ����ӳٻ��䵥�ʱ�/����)��������������ѧϰ����(Auditory Verbal Learning Test, AVLT)�����߾�����Ա�����һ����֪���ճ�����û���ϰ�������û�гմ���
�ų�������������״̬���Է���<24������δ���Ƶ�ҽѧ���;���ʷ��˯���ϰ�����ƻ�����ҩ�MRIɨ���з����Բ����䣻������Ҫ������������ϵͳ��ҩ��(�翹����ҩ��������ҩ������������κ����͵ķǴ�������ҩ�����Ӳ�)���������ĵ������������⣬ͨ��������������II (����÷�>19���ų�)��MRI�ų���Ҫ�Ĵ��Բ�����������������ǰ���з硣���⣬�������������������so-tDCS��ٴ̼�blocks˯��ʱ�䲻��������Ȳ�������ô���Ǿͱ��ų��ڷ���֮�⣬���Ӧ��˯�߽�2����˯��<22���ӡ������������ǵĴ̼��������÷���ּ�ڷ�ֹ�ڲ��ʵ��Ĵ���״̬(���ѡ�˯�ߵ�һ�λ�REM˯��)�ڼ�Ӧ��so-tDCS��
�ܵ���˵�� 6�����ߵ����ݲ��ò����ų���2������˯�߲��㣬3������ɵ��Ի�����1��������ǰδ�����ֵľ����ϰ���������16�����������ʵ�����ų��Ļ��߳����ܽ���ʱ���⣬���߲�����ö�������(�ܽ���ʱ��p=0.048��������p>0.2��������������1)�������쳣ֵ(30��)�����ų������ߵ�ƽ���ܽ���ʱ�������
��1������������
2.2 ��������
��ÿλ�����߽�����ȫ���������ѧ���ԣ���������������֪״̬�������������(�������������ѧϰ����)����������(���ֹ��)��ִ�й���(Stroop��ɫ-���ʲ���)������������(��˹�����������Բ���)���ӹ��ٶȺ�����ת��(�����ƶ����Ե�һ���ֺ͵ڶ�����)��ѡ����ע��ͼ��С����⣬ʹ������CERAD-Plus������������ѧ�����顣����ʱ�����״̬ʹ�û�������������������������������1��
���⣬���й������˯��ϰ�ߵ��ʾ�(�����ij���-����-�ʾ�)��˯�������ʾ�(ƥ�ȱ�˯������ָ��)������˯���ʾ�(Epworth��˯����)�Ͱ�ɭ�����������˯���ʾ����顣ÿ��˯���ռǺͻ��¼��(GT3XActiGraph)���ڼ��ʵ����˯ǰ7���ϰ���Ծ���ʱ�������ʱ�䡣��Щ����֤ʵ�˲�����ֹ��ɵ�˯��ʱ�����
2.3 ����
ʵ�������Ldenbauer����(2016��)��ͬ�������֮����������̼��ͼٴ̼���������ƽ�⽻����ƽ��в���(�ڵ�һ��ʵ������˯ʱn=8�������߽�����so-tDCS��n=8�������˼ٴ̼�)���������2���Է�ֹ����ЧӦ����ʵ������˯֮ǰ����������ʵ�������������Ӧ����˯��
���е���˯���ڵ¹��������ɴ�ѧ��˯��ʵ���ҽ��С�������11:30����ʱ������������EEG��¼��Ȼ����������������Լ�������(��ͷ�������ѧϰ���Ӿ��ռ�ѧϰ������ͼƬ��λ�ü���)������dz����Լ�������(��ָ˳���û�)������2:00����һ�ٱ�����С�ͺ�����so-tDCS�����߱�Ҫ����˯��90���ӣ�Ȼ��������30���Ӻ���м�����ԡ�
��ѧϰ֮ǰ����˯֮��Ļָ�֮ǰ��ע��������(ע�������ֲ���)������״̬(PANAS����)����˯(ƣ��֢״�������Ӿ�ģ������)�ͼ���(���ڽ��ŵ�VAS)�������Կ��ƿ��ܵĻ���ЧӦ��ʵ����̵�ʾ��ͼ��ͼ1a��
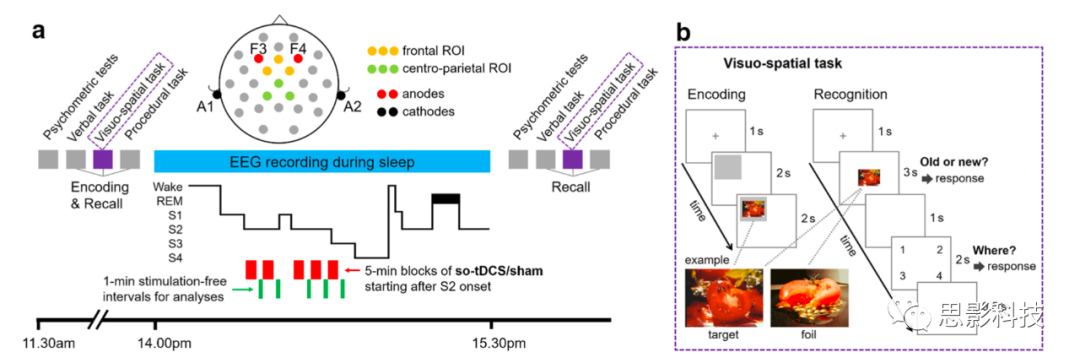
ͼ1 �о���ơ�
a.�������������Ʋ��Ժ������߰���ָʾ��˳��ѧϰ���ԡ��Ӿ��ռ�ͳ�������������90������(����2����3��30��)����¼�Ե�ͼ����Ӧ������ֱ����̼�(so-tDCS)��ٴ̼�(�������ڲ���ƣ����˳��)������˯��30���ӽ��м�����ȡ�������������Ʋ��ԡ�
b.�Ӿ��ռ���������ı����ʶ��ʵ����
2.4 so-tDCS
�̼�������Ladenbauer����(2016��)��������ͬ���̼��缫(ֱ��8����)λ�ڹ���10-20ϵͳ�Ķ�ҶF3��F4����(��װ��EASYñ��)���ο��缫������ÿ����ͻ(ͬ�ࣻͬ��ֱ��Ϊ8����)�����������ɵ�������Ĵ̼���(DC-Stimulator��NeuroConn������������˫��缫λ��)����0.75 Hz��Ƶ��������(��0-262.5��A֮��)������0.522 mA/cm2���������ܶȡ��缫����<5 k����
so-tDCS�������߽����ȶ��ķǿ����۶�(non-rapid eye movement, NREM)˯�ߵڶ��κ�4���ӿ�ʼ��5����һ��block���д̼���(����)1��40����̼�����Ϊblock��ļ����ÿһ��1�����̼������ʼ�ı��(���ڴ̼��յ�ЧӦ����������)������ÿһ���̼����ڽ�����40���ֶ����ã��Դӷ������ų�δ�˲�������EEG�ź��пɼ���ǿ�Һͳ־õĴ̼��յ�Ư��(�̼�����ķ������������г�Ϊ��1�����̼������)����Ladenbauer����(2016��)������5���Ӵ̼�blocks ������(3�C5��blocks������3)���м��̼�����ij���ʱ��ȡ���ڸ��������ߵ�˯�ߣ���Ϊ��ÿ���̼�block֮����˯�ߣ�����ÿ���̼�block����NREM˯�ߵ�2��3��4�ο�ʼ�������������so-tDCS/�ٴ̼�block���˯�߽�2��3��4����˯�߽�1 (��REM˯��)������������״̬�������ӳ����̼������Ŀ�����ڸ����˯�߽�ѡ���Եؽ��д̼�����ÿ���̼�����ڼ䣬��������˯�����֣����ӳ���һ�δ̼���ֱ�����������½���ڶ�˯�߽�1���ӡ���Ϊ��ǰ���о�������so-tDCS��Ӱ����Ҫȡ�������ڽ��еĴ���״̬���÷�����ѡ�����ڽ��������˽ϸߵ�˯����Ƭ���Է�ֹ�ڲ��ʵ��Ĵ���״̬��(����ѡ�˯�ߵ�һ�λ�REM��)Ӧ��so-tDCS��
�ڼٴ̼��ڼ䣬�̼��缫����������so-tDCS��ͬ��λ�ã�����tDCSװ�ñ��ֹرա�����so-tDCS������ͬ�ı�Ӧ���ڼٴ̼�����(��һ���ٴ̼�block�ڵڶ�˯�߽ο�ʼ��4���ӿ�ʼ���������ļٴ̼�block����Ҫ˯�߽�2������˯��)��ͬ����ÿ���ٴ̼�block���ǰ40�뱻�ų��ڷ���֮�⡣�������о������У������߲�֪���̼�������������������о���صij�������DZ��ʼ��Ƿ��ܹ��³����ĸ�ʵ����С˯��ʩ���˴̼����Լ�������С˯���Ƿ����κθо���
2.5 ˯����Ԥ����
����˯�ڼ䣬ʹ�ø�����չ��10-20����EEGϵͳ���õ�Ag-AgCl���ε缫����26��ͷƤ��λ(FP1��FP2��AFz��F7��Fz��F8��FC5��FC1��FC2��FC6��C3��Cz��C4��T7��T8��CP5��CP1��CP2��CP6��P7��P3��Pz��P4��P8��O1��O2)��¼EEG��FCz�������ӵء�ʹ��BrainAmp�Ŵ���ϵͳ(Brain Products)��500 Hz�IJ������ʼ�¼���ݣ�����0.05��127 Hz֮����д�ͨ�˲������еĵ缫��¼���ο��˸����ڱ����ϵĵ缫���迹<5 k�������⣬���ݱ�˯����¼�°͵�EMGs�Լ�ˮƽ�ʹ�ֱEOGs��
��Ӧ���ݲ��˲���(����Ƶ��Ϊ50 Hz������Ϊ5 Hz)��ԭʼ���ݽ����˰��Զ����Ӿ�ɸ�飬��������α����Ⱦ��EEG�Ρ���һ��������BrainVision Analyzer (Brain Products)������ɵġ�
2.6 EEG����
������ÿ��so-tDCS�ͼٴ̼�block���1�����̼�����ڽ�����Ƶ����λ�����Ϸ���������������ʵ�ʽ��д̼���ٴ̼��Ĵ��������������������з�����
���ʡ�ʹ�ÿ��ٸ���Ҷ�任����ÿ���缫ÿ1�����̼������Ƶ���ʣ����11���ص�(5��)��α���Σ�ÿ������10�롣��Ӧ�ļ�����ڼٴ̼����ڼ��㹦����(Ƶ�ʷֱ���Ϊ0.06����)֮ǰ����ÿһ��10���EEG���ݶ�Ӧ�ú����������ƽ������(��V2)���ڸ���Ȥ��Ƶ���ϼ���ģ�SO(0.5-1 Hz)�Ϳ��ٷĴ���(12-15 Hz)�����ǻ����������Ĵ���Ƶ��(8-12 Hz)����������ǰ�Ľ�����бȽϡ�������ǰ���о���ѡ������ЩƵ���ĸ���Ȥ��������(regions of interest, ROIs)���缫λ��FC1��Fz��FC2�Լ�CP1��Cz��CP2�ֱ�ϲ�������ROIs������Ҷ�����붥Ҷ����ˣ�ƽ�����ʷ�ӳ��ÿ��ROI�缫��ƽ��ֵ��ʹ��BrainVision Analyzer�����������ʷ�����
��λ������(Phase�Camplitude coupling, PAC)��Ϊ������EEG�ź����������ص���֮���ʱ���ϵ������ʹ���˻���MATLAB��FieldTrip��������Զ����MATLAB�����������¼�����������������˵������ͨ��Ӧ�����³����Զ�ҶROI (Fz��FC1��FC2)�����붥ҶROI (Cz��CP1��CP2)��SO-to-spindle PAC�����˱�����
(i)�����ѽ����ļ���㷨��Ϊÿ�������ߡ������͵缫ȷ����SO�������¼�:
���ȣ�EEG������0.16��1.25 Hz֮���˲�(˫ͨFIR��ͨ�˲���������=3���ͽ�ֹƵ������)����ʹ����αӰ���ݣ�����״̬��REM˯����Ҳ���ų����¼����֮�⡣
�ڶ�����ѡSO�����ڱ�ȷ��Ϊ�˲��ź������������������������֮���ʱ�䡣ѡ�����SO����ʱ������¼�(�ڼ���0.8 s����2 s����Ӧ��0.5-1.25����)��
������Ϊʣ��ĺ�ѡSOȷ���¼����(���������㽻��֮��IJ���-�������)��Ҳ����SO��������¼�(��SO��ѡ�����75%�����������25%���¼�)����Ϊ��SO�¼�����������¼���δ���˵�ԭʼ�ź�����ȡ���������˲��ź���SO �½�״̬����α����(-2.5��+2.5 s)��
(ii)ʹ�û���(10 ms����) Hanning�δ���(���пɱ��Ƶ����������)����ÿ���¼��κ�ͨ����5-20 Hz (����0.25 Hz)��ʱƵ��ʾ(Time�Cfrequency representations, TFRs) (FieldTrip�������mtmconvol����)�����ڳ����������������������ɣ���ȷ���ɿ��Ĺ��ʹ��ơ�Ȼ��TFRs����Ϊ�¼�ǰ���ߵIJ�ֵ(EEG�ε�-2.5��-1.2��)�����������ߡ�������ROI����ƽ����
(iii)Ϊ���������ٷĴ������ʵ�(���ڵ��Ƶ�)SO�� (�����˵�) ����֮���ͬ�������������Ǽ�������Щʱ�����е���λֵ����Ӧ����ͬ��ָ����ʹ��ϣ�����ر任Ϊÿ����ȡ��SO�¼�������ʱ������Ӧ�Ŀ��ٷĴ������ʲ���������λֵ���Ĵ������ʲ���ʱ������ͨ���ڸ���Ƶ����ƽ����TFR���ã������ϲ�����500���ȵIJ���Ƶ�ʡ�Ϊ��ȷ����ȷ����λ���ƣ��ڵ���SO�¼�(0.5-1.25���ȣ�˫ͨFIR��ͨ�˲���������=3���ͽ�ֹƵ������)��Ȼ�����ÿ���¼��κ͵缫��������λֵʱ������֮���ͬ��ָ��(synchronization index, SI)�����õ�SI�Ǹ�������뾶(r)��ʾ����SO�͵������ٷĴ������ʲ���֮�������ǿ�ȣ��Ƕ�(��)��ʾ��Щ��֮������ơ����仰˵������ʾ���ٷĴ�������ʱ��仯����SO�Ρ�����ͨ�����·�ʽ��õģ�
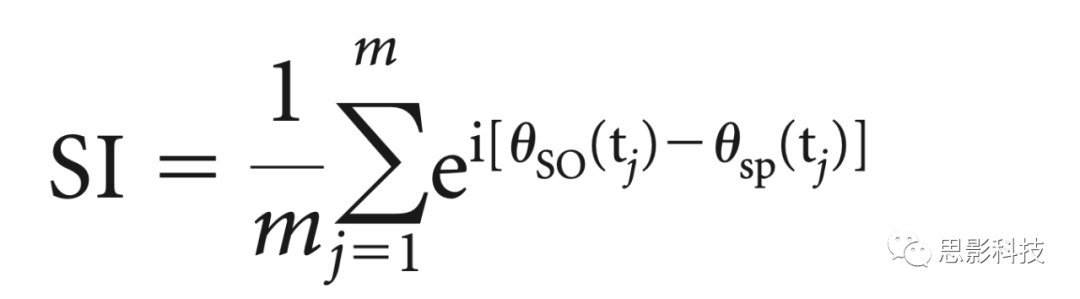
����m��ʱ����������SO(tj)��ʱ���tjʱSOʱ�����е���λֵ��������sp(tj)��ʱ���tj�����ٷĴ�����ʱ�����в�������λֵ������SI��ʱ������SO���ĵ�-1 s��1 s��ÿ��������ROI��SIs�ֲ���ͼ3d�п��ӻ�������ͳ�Ʒ�����SIs�������ߡ�������ROI����ƽ����

ͼ3 SO��Ĵ�������֮�����λ�����ϡ�
a. EEGƽ��ֵ��
b. TFRs������SO�¼���ƽ��ÿ��������
c.�ڿ�Ĵ���(12-15Hz����)�����Ĵ���(8-12Hz����)Ƶ�ʷ�Χ�ĵ���SO��Χ�ڣ����˲����TFRs���¼�����ƽ�����ʵ�ʱ����̡�
d. SI��ֱ��ͼ��
e.���������ÿ��������(ϸ��)�Ϳ�������(����)��SIsƽ��ֵ��
˯�߽ṹ��˯�߽ṹ�������ڲ�ͬ˯�߽λ��ѵ�ʱ��ͱ������ǻ��ڶർ˯��ͼ��ȷ���ġ�Ϊ�ˣ�EEG���ݱ��²�����250���ȣ�����˯�߽�1��2��3��4�Լ�REM˯�ߡ������ڻ��˶�αӰ�У�ͨ��Schlafaus����(Steffen Gais)�ֶ�����30���EEG�Ρ�����EEG�ź��е�����αӰ��so-tDCS�ڼ�Ķ�û�����֡�ͬ�����ٴ̼��е���Ӧʱ��Ҳû�����֣��Ի�ÿɱȽϵ�˯��ʱ��ͱ�������10���ʱ����ڣ���1�����̼����(���Դ̼���5����blocks֮��)�������֡�
2.7 ��������
���м�����������Presentation�������У�ƽ�а汾����������ʵ������˯�ε�����������Ladenbauer����(2016��)�У����м�������Ҳ��ִ�й������Ӿ��ռ�����Լ��������У������߱�Ҫ�������Ŀ�Ա��պ���䣬��û���Ƽ�����IJ��ԡ��Ӿ��ռ�����Լ���������ʹ�õĴ̼�û���ص���
���ǽ��Ӿ�ʶ�������Ϊ��Ҫ����Ϊ������������Ϊ����AD�����ܵ�Ӱ�죬��/��ʶ����������ɿ���Ԥ�ⲻͬ�� (AD��MCI����������)����������䷶���෴��ʶ���������������Ŀ��������������ȷ�������Ŀ��Ϊ������ǰ���о����бȽϣ����Ƕ���������λ�û��������ʶԻ��䡣
�Ӿ��ռ���������Ӿ��ռ����������38������ͼƬ(���Թ������ͼƬϵͳ�����塢ֲ������ͷ羰)��ɣ�����Ļ��4�����ܵ������е�1���������2���ӣ����1����(��ͼ1b)��Ϊ�˽�������ЧӦ�ͽ���ЧӦ���ڷ����м����������ķ�ͼƬ(�����ڿ�ʼ�������ڽ���)�������������ǡ������߱�Ҫ���סͼƬ(ͼƬ����)�����ǵ�λ��(λ�ü���)����ʶ����Թ�����(˯��ǰ��)��ÿ��ͼƬ(38���о�����ͼƬ��38��������е���ͼƬ)��ʾ����Ļ����(3����)��ͬʱ������ͨ�����°�ťָʾ�����Ƿ�ʶ����ÿ��ͼƬ(�DZ�ʾ���ɡ����ߣ����ʾ���¡�����)�������������ʶ��һ����Ŀ(���ɡ�����)����ô���ǻ���Ҫָ��������ΪͼƬ���ڲɼ������г������ĸ����ޡ�
�������е�����/�������ߵ������ֿ��ܵ���Ӧ�������(��ȷ�����ɡ��ж�)����ȷ�ľܾ�(��ȷ�����¡��ж�)������ľ���(����ȷ�����ɡ��ж�)�ʹ���(����ȷ�����¡��ж�)����ΪͼƬʶ�����ȷ�Եĺ���������ȷ�ش�İٷֱȼ������£����б���+��ȷ�ܾ��ı��������⣬ͨ���������к��ı���֮�ͣ�������DZ�ڵķ�Ӧƫ�
Ϊ��λ�ü����ȷ�ԣ���ȷ�ʹ��������ͼƬλ�ö������Dz��������£���ȷ������λ����/������-���������λ����/�����������ڼ������⣬һ�������ߵ����ݱ��ų��ڱ�����ķ���֮�⡣
���ǵ�����ͼƬ�̼�(�����ͷ羰)�ĸ߱����Լ�Ŀ��ͳ��е������ԣ�ͼƬʶ����ְ����������Ϥ���̣������ֶ��ѱ�֤����MCI���ܵ�Ӱ�죬��˾����ٴ�����ԡ�
��ͷ�������������߹ۿ���40��������صĵ��ﵥ�ʶ�(���-ʵ���ԣ����磬ˮ��-�㽶)����Щ���ʶ�����Ļ���������5���ӣ��м���Ϊ100���롣����������ĸ����ʶ�(�����ڿ�ͷ�������ڽ�β)���Է�ֹ����-����ЧӦ��������г�ʼ����������ԡ��������(����)����������λ�ã���������ָʾ(10���ڰ��°�ť)�Ƿ��ס���ԵĴ̼�(ʵ��)�ʺ�Ҫ�����˵�����ԵĴ̼�(ʵ��)�ʡ������ȷ�ĵ��ʶ���ʾ��2.5�롣�����ʼ����-��������ṩ��һ�������ѧϰ���ᣬʹ��������������������У�~60%�Ļش���ȷ�������������-������ӳ�����-�������(˯�ߺ�)��û�ж��������ȷ���ʶԵ�����½��С����ʶ��Բ�ͬ�����˳����֣��Է�ֹ��ÿ������ͻ�������е�����ѧϰ����¼���Ե����ﷴӦ�������ݼ�ʱ������ӳٻ�������ȷĿ��ı����ֱ�����������ɼ���
���⣬���Dz����˴̼��Ը����ʵ�Ӱ��(��˯ʱ���ж�ǰһʱ����ͬ������ȷ��Ӧ�IJ���ȷʵ���ʵı���)������һ�������ߵ����ݱ��ų��ڸ�����ķ���֮�⣬��Ϊ��δ����ѭ����˵����
��ע�⣬���ǵĿ�ͷ����ͬ��Marshall����(2006)��Τ˹�ر������(2015)��ʹ�õ�������Ϊ���ǵĴʶ��б�ּ��ʵ������ʵ����С˯�Ĵʶ�֮��ĸ߶ȿɱ��ԣ����ų���дʡ����ܴʶ��б�֮��Ŀɱ��������-ʵ��(���磬ˮ��-�㽶)�ṹ�����ӣ�������ǰ���о���ȣ��ʶԱ��ֳ���ǿ�����������
����������һ���ı���Walker����(2002)��������ָ�û����������о�so-tDCS�Գ�������Ӱ�졣�����߱�Ҫ����30���ʱ�����ڣ��Ծ�����ȷ�Ϳ��ٵķ�ʽ�����û���������������Ļ����ʾ����λ������(����4-2-3-1-4)(����)������˯ǰ����(ѧϰ)�У������߽�����12�β��ԣ��м�30����Ϣ�������˯�����(����)�����Ĵ�����Ϣ�����顣ѧϰ�ͼ������Եı��ֱַ���������������е�ƽ����ȷ�û����о���������������δ���������������˱��ų�����������ķ���֮�⡣
2.8 ͳ�Ʒ���
���ʡ�ʹ�þ�������ؾ�����Ի��ģ��(linear mixed model, LMM)���������ֽ������(����Ҷ�����붥ҶSO���Ĵ����ʣ��Լ���Ҷ�Ĵ�����)�����ִ̼�����(so-tDCS�ͼٴ̼�)֮��IJ��졣ѡ�����ģ����Ϊ�˽���˯������ѧ�е��͵ĸ�������Լ���ͬ�����ߺ������µIJ���������Ƶ�������ݵ�(���ݸ��������ߵ�˯�������Ӧ���������so-tDCS�ͼٴ̼�blocks)����Щ��ƽ���������ڲ��ʵ��Ĵ���״̬�²�����so-tDCS��
�����֮��ģ���������£�
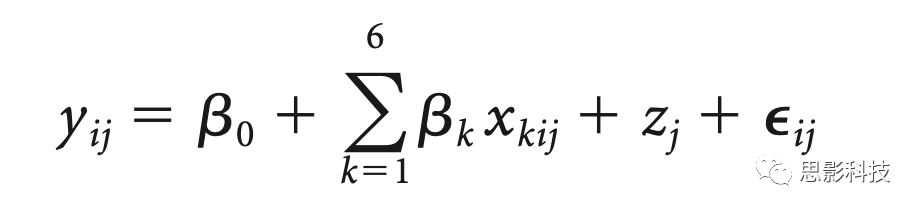
����yijָ������j��{1,2,��,16}�ڼ��i��{1,2,3,4,5}��log�任�������ֵ����0�ǹ̶��ؾ࣬��1 ,��, ��6�Ƕ�Ӧ�ڴ̼��Ա����Ļع�ϵ����x1ij��{0,1}�����ߣ�x2ij��(����)ʱ�䣬x3ij=i-3��(����)ʱ���ƽ����x4ij=(i-3)2��ʱ�����̼���x5ij=(i-3) x2ij��ʱ���ƽ�����̼���x6ij=(i-3)2 x2ij���ض����Ե�����ؾ���zj�����ģ���ij�Dzв�(��������)��Ҳ����˵�����ʱ���(ÿ���̼�block��1�����̼����)��Ƕ�����������е�һ��ˮƽ��Ԫ(����ˮƽ��Ԫ)����ģ�ͼ���б�������Ƶ�(û�����б��ģ��)����Ϊ֮ǰû��֤�ݱ����̼�ЧӦ(б��)��˯������ѧ�ĸ������졣��ΪЭ�����Ļ��߱���(x2ij)�ֱ����ڵ���ÿ��Ƶ���Ļ��߲��졣�ñ������ôӵ�һ�δ̼�/�ٴ̼�blockǰ��1���Ӽ������������������Զ����任Ƶ����ֵ�������������ݲ�������̬�ֲ��������Ӧ��LMM֮ǰ��������˶����任����ע�⣬�̼��ͻ��߱�����ֵ�ڲ�ͬʱ������û�б仯��ƽ������ʱ�����(x4ij)���ڲ���SO�ͷĴ���������ʱ���������߽��̡�ͨ�������ʱ�����̼������������˴̼������������ʱ���ƽ�����̼�֮�������б���Ƿ�ͬ�������̼�����֮���������״�Ƿ�ͬ��
PAC����λ��������������ʹ��˫β�������t����(��ˮƽͳ��)�����˴̼������TFR���������¼��������ʱ仯��Ϊ��У�����رȽ�(-1.2s��+1.2 s �� 5-20����)����FieldTrip��Ӧ����һ�����ھ�����û��������ſ鶨��ij�ʼ��ֵ����Ϊp<0.025���������ܺ�tֵ�����Ե�������ֵ����Ϊp<0.05��
Ϊ������(Բ��)SI���Ƿ����ض���ƽ��ֵ�����ȷֲ�������Ӧ����V���顣ʹ��������ԣ��������H1ָ�����˿ھۼ���һ����֪��ƽ�������ڵ�ǰ����£�����������SO����״̬�������������ٷĴ������ʣ�Ҳ����˵���ۼ���0�����ҡ�
���⣬���Ǽ����˽���������ȣ���������SI�ǵļ��г̶ȡ������ض���������ROI�����㷽�����£�

������j�DZ���j��{1,2,��,n}��SI�ǡ�
˯�߽ṹ��Ϊ������so-tDCS��˯�߽ṹ��Ӱ�죬��������˯�ڼ��1�����̼�����ڼ䲻ͬ˯�߽������ѵ�ʱ�����(��λΪ%)������t����(��Wilcoxon�����Ȳ���)��
��������ͨ���ظ������������(repeated-measures ANOVA, rmANOVA)������so-tDCS�Լ��������б��ֱ��ֵ�Ӱ�죬���������������ش̼�(so-tDCS�ͼٴ̼�)��ʱ���(��˯ǰ����˯��)���ʵ�ʱӦ�������ɶȵ�Greenhouse-GeisserУ����
������У������Υ�������μ�������ʱ����Ҫ����epsilon(��)У������ epsilon(��)�ӽ� 1 ʱ��Greenhouse-Geisser У�������ڵ� epsilon(��)��������У������ Huynd-Feldt У�������ڸ߹� epsilon(��)���������ɵ�������ͨ��������ʹ�� Greenhouse-Geisser У���������ǵ����Ƶ� epsilon(��) С�� 0.75 ʱ�����ǣ�������Ƶ� epsilon(��) ���� 0.75��һЩͳ��ѧ�ҽ���ʹ�� Huynd-Feldt У������ʵ���У�����������������dz����Ƶ����������������Ƶ� epsilon(��) ���� 0.75������ʹ���κ�һ�֡���
ʹ��SPSS���м��������ʡ�˯�߽ṹ��ͳ�Ʒ�����ʹ��FieldTrip��CircStat MATLAB�������Լ��Զ���MATLAB������PAC��Ϊ����ͳ�Ʒ�����˫��������ˮƽ�������з����б�����Ϊ0.05����������Ȥ����Ҫ�����Ķ������[n=5���Ӿ�ʶ�����(��ȷ�����ٷֱ�)�������������ص�EEGƵ���ʲ���������Ҷ�����붥ҶSO�Ϳ��ٷĴ�������]�����Dz���Benjamini-HochbergУ����������q=0.05�Ĵ������ʡ�У�����������ˮƽ�ڽ��������q*��ʾ��Ϊ��У��ͳ��TFR�����еĶ��رȽϣ�Ӧ���˻��ھ�����û���������ΪЧӦ�ı���ЧӦ��С��ƫ��2(�������Ϊ��2)�����⣬�������ñʾ�ֵ������˴̼���EEG����Ӱ���LMM�����Ի��ģ�ͣ���ģ�ͱ���ЧӦ��С(Cohen��s d)��������SEs������SD��������������ͱȽ϶����Ҫ�����йأ�pֵӦ����̽���Է����Ŀ���ڽ��н��͡�
��������Ե�����ݴ�������Ȥ����ӭ���˼Ӱ�Ƽ���վ�����ں��˽���ص������γ̡��������������������������ź�siyingyxf��18983979082��ѯ����˼Ӱ�ṩ����������ط�������ҪҲ�����Ӵ��ź���Ⱥ)��
�Ͼ���
�ڶ�ʮ�����Ե����ݴ������Űࣨ�Ͼ���4.23-27��
������
�ڶ�ʮ����Ե����ݴ������Ű�(������5.16-20��
�Ϻ���
�������Ե����ѧϰ���ݴ����ࣨMatlab�棬�Ϻ���5.15-20��
����ʮ�����Ե����ݴ����м���(�Ϻ���5.28-6.1��
���죺
�ڶ�ʮ����������Թ������ݴ����ࣨ���죬5.24-29��
���ݴ���ҵ����ܣ�
˼Ӱ�Ƽ�EEG/ERP���ݴ���ҵ��
˼Ӱ�Ƽ��������Թ������ݴ�������
˼Ӱ�Ƽ��Ե����ѧϰ���ݴ���ҵ��
˼Ӱ���ݴ������������Դ�ͼ��MEG�����ݴ���
˼Ӱ�Ƽ��۶����ݴ�������
��Ƹ����Ʒ��
˼Ӱ�Ƽ���Ƹ���ݴ�������ʦ ���Ϻ����������Ͼ���
BIOSEMI�Ե�ϵͳ����
Ŀ��ʽ���ܴŹ���̼�ϵͳ����
3.���
���Ƿ�����16��MCI����(7��Ů�ԣ�ƽ������70.6����8.9 SD)����������˯90����ǰ������˼���������ԣ�������ʹ����so-tDCS��ٴ̼�(���Σ�ƽ�⽻�����)��ʵ���ʾ��ͼ��ͼ1��ʾ������רע���Ӿ�ʶ����䣬�������˷�����AD�������ڣ���˹���MCI�������Ƶ����ае㡣Ϊ������ǰ���о����бȽϣ����Ƕ��������˴ʶԻ��䡣
��������ѯ��so-tDCS�Ƿ���������ص�˯�߲����������Ӱ�졣Ϊ�ˣ������ص��о����ڳ���5��so-tDCS/�ٴ̼�blocks�����1���ӵ��̼�����ڼ䣬ͨ��EEG�����Ķ�SO�Ϳ��ٷĴ����ļ�ʱӰ�졣������������Щ�Ĺ����Լ����ǵĹ�����ϡ�������ǰ���о������Ƿֱ�Զ�����붥Ҷ��ROIs�����˷��������⣬���������˶�Ҷ���Ĵ������ʣ���������˯�߽ṹ�仯��Ȼ�������о��˴̼���θı�����Լ���ı��֣����̽���˱��ֵı仯��˯�߲���֮��Ĺ�ϵ��
so-tDCS/�ٴ̼�blocks�������ڲ�ͬ���Ƴ�֮����ܻ�������ͬ����Ϊ����ͨ��ȷ��ÿ���̼�block֮ǰ����˯�߽�2����������Ǹ��������ߵ�˯��(״̬�����Ĵ̼�)���ڲ����߽��ܵ�so-tDCS/�ٴ̼�blocks���������棬û�й۲쵽��������(����2��Wilcoxon�����ȼ��飺p=0.453)�����л��߶����ִ̼����������á�����û�б����κθо�������һ�����߱�ʾ�д�ʹ�С�ʵ���Ļ㱨��������������������³����ĸ���˯ʱ��Ӧ����so-tDCS (n=14�˻ش�����֪������2����������ȷ�µ���so-tDCS��˯)�����ǵ�ʵ��ǰ��˯��ʱ�����˯�߳���ʱ�䣬�̼�����֮��û�����Բ���(����p>0.2)��
��2 so-tDCS/�ٴ̼����������˯�߽ṹ
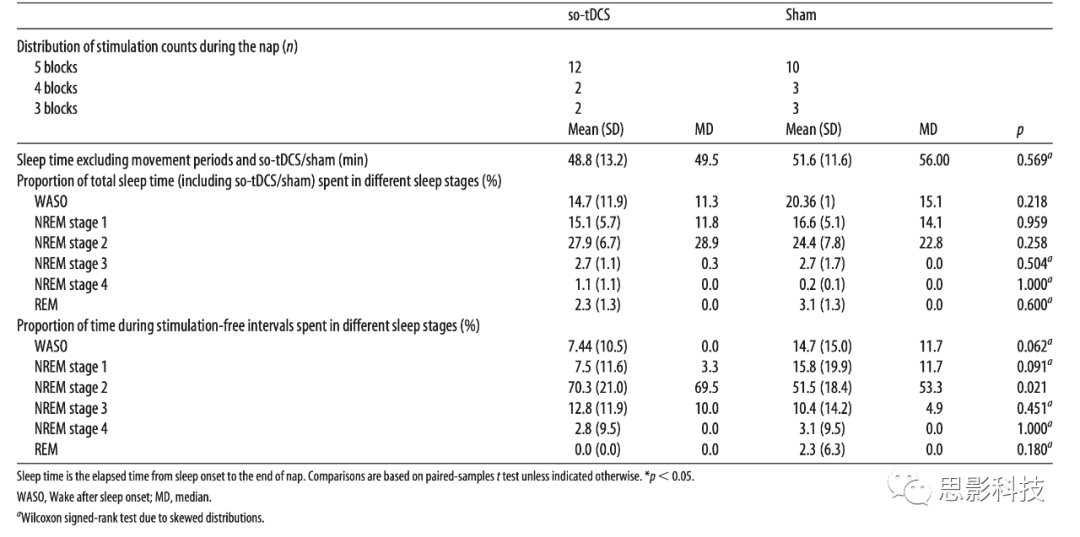
3.1 �Լ������˯�߲�����Ӱ��
3.1.1 Ƶ����
Ϊ�˷����̼���Ƶ���ʼ������Ƶ����Ӱ�죬����ʹ����LMM�����Ի��ģ�ͣ��������÷���������˯������ѧ�еĸ�������������֮�䲻��ȵĹ۲����(3-5��so-tDCS/�ٴ̼�blocks)��LMM������ʾ������˯�ڼ䣬��Ҷ�����붥Ҷ��SO (0.5-1 Hz)������������so-tDCS�����ǿ[�̼���ƽ�����죬��Ҷ��0.28��95%��������(confidence interval, CI)��0.16-0.40��d=1.16��p<0.001��q1=0.01�����붥Ҷ��0.19��95% CI��0.06-0.31��d=0.70��p=0.006��q3=0.03��ͼ2)��������ǿ�����û���SO����(�ڵ�һ��so-tDCS/�ٴ̼�blockǰ1������)�����ͣ��ڸû���so-tDCS�ͼٴ̼�����֮��û�й۲쵽��������(��Ҷ��t(15)=1.65��p=0.121�����붥Ҷ��t(15)=1.20��p=0.249)�����⣬����SO������̼�blocks���SO������������(���ߣ���Ҷ����2=-0.082��SE=0.168��p=0.628�����붥Ҷ����2=-0.145��SE=0.206��p=0.483)������˯���ڼ�1�����̼�ʱ������SO���ʵĹ��̣��������أ�����û�й۲쵽���������Ա仯(ʱ�䣬��Ҷ��3=0.027��SE=0.029��p=0.353�����붥Ҷ����3=0.027��SE=0.033��p=0.408)�����ʵ�U��ϵ�����ֹ�ϵ�����붥Ҷ��ǿ(ʱ���ƽ������4=-0.061��SE=0.027��p=0.025)�ȶ�Ҷ��λ(ʱ���ƽ������4=-0.040��SE=0.024��p=0.092)���ڲ�ͬ������(ʱ�����̼���ʱ���ƽ�����̼�����ã�����p>0.2)��
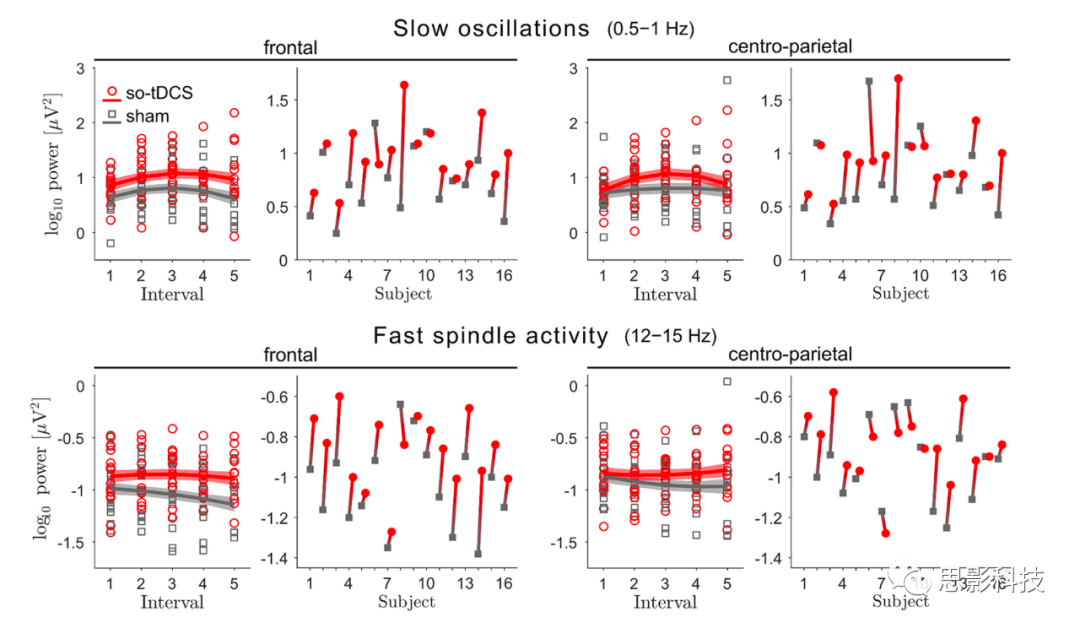
ͼ2 so-tDCS�����SO�Ϳ��ٷĴ���Ƶ�ʷ�Χ�ڵ�EEG����
������ǿ��so-tDCSЧӦ����������so���Ρ����ڿ��ٷĴ�������(12-15Hz)��LMM����Ҳ��ʾ�������Ĵ̼�ЧӦ(�̼���ƽ�����죬��Ҷ��0.14��95% CI��0.07-0.21��d=1.02��p<0.001��q2=0.02�����붥Ҷ��0.07��95% CI:0.003-0.14��d=0.50��p=0.041��q5=0.05)������ٴ̼���ȣ�so-tDCS��Ĺ������ӡ���Щso-tDCS�յ��Ĺ�������Ҳ�����û����ڼ�Ŀ��ٷĴ������������ͣ���Ϊ�ڻ����ڼ䣬so-tDCS�ͼٴ̼�֮��û����������(��Ҷt(15)=1.84��p=0.086�����붥Ҷ��t(15)=0.88��p=0.393)��Ȼ�������Ƿ��ֻ��߿��ٷĴ����������Ƶ�ʷ�Χ�Ĺ��������Ĵ̼����ɼ���ڼ��Ҷ(���ߣ���2=0.463,SE=0.122,p<0.001)�����붥Ҷ(���ߣ���2=0.392,SE=0.146,p=0.010)������ء�
���ǽ�һ���о��˴̼��Զ�Ҷ���ٷĴ�������(8-12Hz)��Ӱ�죬��Ϊ֮ǰ�������������е�һ���о�����so-tDCS�յ������Ĵ��������ӣ����Կ�Ĵ�������û��Ӱ�졣�ڱ��о��У���ٴ̼���ȣ�so-tDCS���Ҷ���Ĵ���������������(�̼���ƽ�����죬0.10��95% CI: 0.04-0.16��d=0.82��p=0.001)��������߶�Ҷ�Ĵ���������so-tDCS�ͼٴ̼�����֮��û����������(t(15)=1.11��p=0.287)�����ǣ����Ĵ�������һ�£�so-tDCS/�ٴ̼�block���̼�����ڼ�������Ĵ����������Ƶ���µĹ����������(���ߣ���2=0.674��SE=0.112��p<0.001)���ڿ�Ĵ���Ƶ�ʷ�Χ�����Ĵ���Ƶ�ʷ�Χ�ڣ�û�з��ֶԹ��ʵĽ�һ������Ӱ��(����p>0.1)��
3.1.2 PAC
Ϊ������SO�������ͷĴ����Ĺ�����ϣ����Dz�����һ�ַ����������о��Ĵ��������(����)�Ƿ�SO����λ(��λ-������)ϵͳ�ص��ơ�ʹ���¼����������������ر����˼����SO�Ϳ��ٷĴ����Ƕ�ס����Ǹ����ѽ����ļ���㷨ʶ����EEG�е�SO�¼��������¼��ε�TFRs��SO���ȵ����Ķ��롣����Ӧ��(�¼�������)TFR�У�SO�ͷĴ�����PAC��Ϊ���ʵ�����ʱ����֡�Ϊ��ȷ���ɿ���PAC��������ò������������ų��ڸ÷���֮�⣬��Ϊ���ǽ����ֳ���������SO�¼�(ÿ���缫<10��)����ˣ����շ�������12����
���Ƿ��֣������¼�Ϊ���ĵ�SO����֮���SO������(�½���)��so-tDCS�������Ķ�Ҷ��Ĵ��������������ӣ���SO����֮ǰ��SO�����Σ����Ķ�Ҷ���Ĵ�������Ҳ��������(ͼ3a-c)������ע�����so-tDCS�����£����Ĵ������ʵĵ��Ƶ��������Ը�ǿ������Ϊ������ڿ��ٷĴ�������������λ�ӳ���~250����(ͼ3c)���ڶ�ҶROI������û�й۲쵽�̼��������ַ�ʽ������PAC������Ӱ�졣
����ʹ��SI���¼�����ķ�ʽ��һ���о���SO�Ϳ��ٷĴ���֮���������ϣ��������˵���SO�͵����Ĵ������ʲ���֮�������������ǿ�ȡ�ͼ3d��e��ʾ��SIsǿ�����ڴ̼�����ļ���ļ������PAC����ǿ��SI�ǵ�����ֲ�ָʾ��SO����λ���ڸ���λ����ѡ���ֿ��ٷĴ������������(ͼ3d)��ȣ����ֲַ���ʾ����ǿ�Ľӽ�����(��λͬ��)�Ĵ̼���ֵ������ͨ���ϳ�ʸ������(so-tDCS��0.20���ٴ̼���0.16)���������У����Ҹ��������SI���ڵ�������(270-360�㣬so-tDCS:35%���ٴ̼�:29%)�������Ĵ������ʷ�ֵ��ѡ������SO�ĺ��������Ρ�������ƽ��SI�ǵ�ͳ������֤ʵ����һ��(ͼ3e)������12������8��(so-tDCS)��5��(�ٴ̼�)λ�ڵ������ޣ���ϳ���������(��ʾ�����ߵļ��г̶�)Ϊ0.89(so-tDCS)��0.58(�ٴ̼�)��V��������һ�����������ڴ̼�������������ƽ��SI�ǻ����ϸ����ܵؼ����ڽӽ�0���λ��(so-tDCS: V=10.16��p<0.00002���ٴ̼�:V=6.84��p<0.003)��
���ڶ�ҶROI�����ǹ۲쵽���Ƶ�(��������)�̼���SO�Ϳ��ٷĴ���PAC��ǿ����������ƽ��SI��(so-tDCS: V=8.50��p<0.0003���ٴ̼�:V=5.67��p=0.01)����Ӧ�ĺϳ���������(so-tDCS: 0.71���ٴ̼�: 0.52)��
3.1.3 ˯�߽�
��2�ܽ�����˯��1�����̼�����ڼ䲻ͬ˯�߽ε���˯��ʱ�䡣����˯��ʱ��Ͳ�ͬ˯�߽λ��ѵ�ʱ�䷽�棬û�з�������֮�����������(����p>0.2).Ȼ��������1���ӵ��̼�����������Ƚ���NREM˯�߽�2�в�������������(t(15)=2.57��p=0.021��ƽ������18.86��95% CI:3.22-34.51)����ٴ̼���ȣ�˯�ߵ�2����so-tDCS���������ӣ���˯�߿�ʼ��Ļ���ʱ���NREM˯�ߵ�1�γ������Լ���(Wilcoxon�����ȼ���:p=0.062��p=0.091)��
����������MCI���ߵ�so-tDCS����������SO���ʣ���ǿ�˿�����Ĵ���Ƶ�ʷ�Χ�ڵĹ��ʡ�PAC������һ����ʾ��so-tDCS����SO�Ϳ��ٷĴ�������֮���ǿ��ͬ�����ر�����SO�����κ������ӷĴ������ʡ���Щ�����so-tDCSЧӦҲ��ӳ��1�����̼�����ڼ��˯�߽ṹ�У����������ȣ�˯�߽�2���ӣ�˯�߽�1�;��ѽγʼ������ơ�
3.2 �Լ����Ӱ��
Ϊ�˿��ƶԼ�����ֵ�DZ�ڻ���Ӱ�죬������������˯ǰ�����ұ���������������˯��ע��������˯ǰ������������(p>0.1)��ͬ��������Ҳû�з���ע�����������ͼ���Ĵ̼������Ա仯(p>0.2)�������ұ�������������ӵIJ��������ƣ�����so-tDCS��˯��ƣ��֢״����(��������)ָ����ʾ(F(1,15)=3.5, p=0.082)��
���Dz�����so-tDCSЧӦ�Ƿ�Ҳ��ӳ��MCI���ߵ��Ӿ����������(n=15)������(���)so-tDCS����˯��Ӱ�죬���ǽ���˯��ΪЭ����������Եļ�����ַ���(ANCOVA)������ͼƬʶ��ȷ��(��ȷ�ٷֱ�:������+��ȷ�ܾ���)�����Ƿ���so-tDCS�����Ӿ����������ЧӦ(�̼���ʱ��㽻����F(1,12)=5.34��p=0.039��q4=0.04;������˯�ڼ�ı��ֱ仯����λΪ%��so-tDCS��2.89��95% CI:0.24-5.54���ٴ̼���-0.96��95% CI:-5.10-3.18, ��2=0.308;��˯ǰ����˯��ķ���Ҳ��ͼ4)�����ǽ�һ�������˲�ͬ��������˯ǰ�IJ���Ϳ��ܵķ�Ӧƫ���˯ǰ���Ӿ�ʶ������ڲ�ͬ�Ƴ�û�в���(t(14)=0.38��p=0.712)��û�й۲쵽����ЧӦ(��˯ǰ��ȷ�ٷֱ�:so-tDCS��83.86��95% CI:78.9-88.8���ٴ̼���82.94��95% CI:73.6-91.5��Ȼ����һ�������ڼٴ̼������´ﵽ100%)�����ڷ�Ӧƫ��������ʺ�����֮����ʾ���������������ȣ�ʹ��so-tDCS����˯��ķ�Ӧ��̫����(F(1��12)=11.83��p=0.005, ��2=0.496)��Ȼ����ͨ���������ʺ����ʷֱ����so-tDCSЧӦ������ϸ�ؼ������ЧӦ��������ЧӦ���������ʷ�������(F(1,12)=14.55, p=0.002, ��2=0.548)�����ʲ�����(F(1,12)=2.54, p=0.137, ��2=0.175)�����⣬�����ڷ�����������so-tDCS���Ӿ������Ӱ�죬��û��У��˯��(rmANOVA)���˴����۲쵽��ٴ̼���ȣ���so-tDCS��ı��������������ƣ���δ�ܴﵽ����ˮƽ (F(1,14)=3.58��p=0.079, ��2=0.204)�����ڴ̼���ʱ�����Ӿ�ʶ����ֵ���ҪӰ�죬û�����Ե�����Ӱ�죬���dz�����������ЧӦ������(�̼���F(1,12)=3.80��p=0.075)��
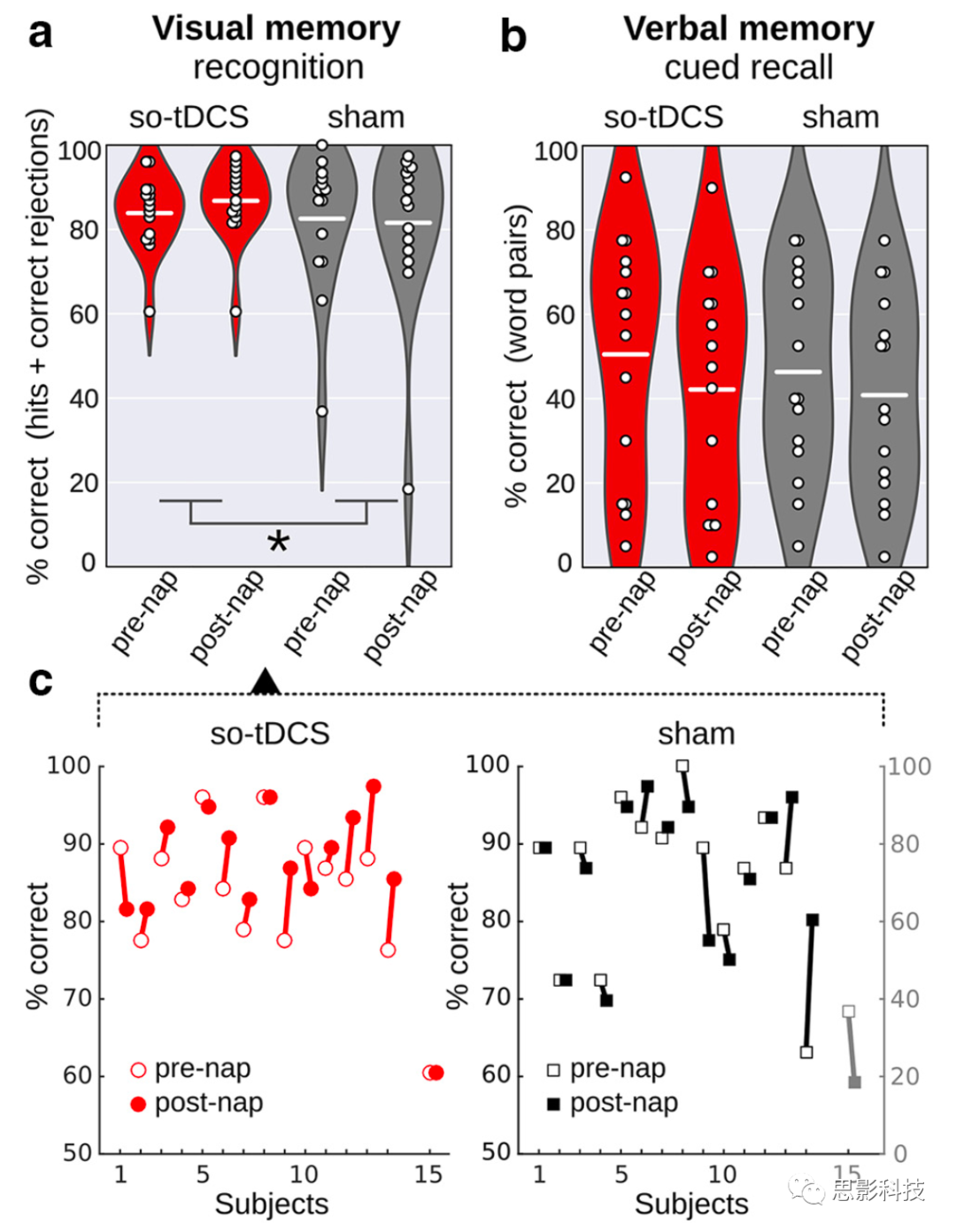
ͼ4 so-tDCS��sham�����������Լ�������ı������ܡ�
a, b����so-tDCS(��ɫ)��sham(��ɫ)����˯ǰ����˯����ͼ�����������(a)�е�ʶ����ֺ������������(b)�е���ʾ������֡�
c, so-tDCS(��)��sham(��)�����¸�����˯ǰ���ͼ��ʶ����֡�
λ�ü������ֲ���so-tDCS��Ӱ��(�̼���ʱ��㽻����F(1,12)=0.01��p=0.913, ��2=0.001;������˯�ڼ�ı��ֱ仯����λΪ%��so-tDCS��8.37��95% CI:-16.62��-0.12;�ٴ̼���-4.95��95% CI:-16.55��6.65)��ֻ��������̼������ص���˯��ռ������½�(��˯ǰ��ļ���仯����ЧӦʱ���:F(1,12)=6.18��p=0.029;��˯ǰ�ٷֱȣ�-12.87��95% CI:-35.84��10.10����˯��ٷֱȣ�-19.53��95% CI:-39.50��0.44)��
Ϊ������ǰ���о����бȽϣ����ǻ��о���so-tDCS�Ե���������ѧϰ����(���Լ���)�м���ɼ���Ӱ�졣����û�й۲쵽so-tDCS��ʾ������ֵ�Ӱ��(������������˯���̼���ʱ��㽻��:F(1,12)=2.27��p=0.156, ��2=0.149��n=15)����MCI��������˯ǰ����˯����ֳ����ԵĶ����ڴ̼������Ļ��������½�(��ЧӦʱ���:F(1,12)=12.96��p=0.003;��˯ǰ%: 45.9��6.6;��˯��%: 39.1��6.1)����˯ǰ�Ļ������Լ��������so-tDCS�ͼٴ̼�֮��û�в���(t(14)=1.03��p=0.322)����û�й۲쵽����ЧӦ(ƽ����˯ǰ��ȷ�÷ְٷֱ�:so-tDCS��50.5��95% CI:35.4-65.6���ٴ̼���46.3��95% CI:33.0-59.7)��
Ϊ��ȷ��so-tDCSЧӦ�������ԣ����˳����Լ��������⣬�������˳����Լ�������û��ע���ЧӦ���ЧӦ��Ӱ��(����p>0.5)��
��֮����У���˻��ӱ�����˯֮��so-tDCS����Ϊˮƽ��Ӱ���ʾ�˶�ͼƬʶ����ֵ�����Ӱ�졣��λ�ü������������Լ��������������������û�й۲쵽so-tDCS��Ӱ�졣
3.3 ˯�߲����ͼ������֮��Ĺ�ϵ
Ϊ���о��̼��շ���˯�߲����仯(SO�ͷĴ������ʱ仯�Լ�˯�߽�)���Ӿ�������ֱ仯֮��Ĺ�ϵ�����ǽ��̼������µ�˯�߲���������so-tDCS�ͼٴ̼������µ��Ӿ�ʶ��(��˯ǰ����˯��)���ֱ仯�IJ�����ϵ���������ڹ��ʲ�����˯�߽Σ�û�з��������������(����p>0.3)��
��������о������Ķ�ҶSO�Ϳ��ٷĴ���֮������(��SI����)�Ƿ����Ӿ�������ֵı仯�йء����Ƿ���so-tDCS��������������˯��-��˯ǰ����������ֲ���������ͬ����SO�Ϳ��ٷĴ���PAC���(SI�ǽӽ���0)�����ڼٴ̼�������û�й۲쵽���ֹ�ϵ(ͼ5a)��
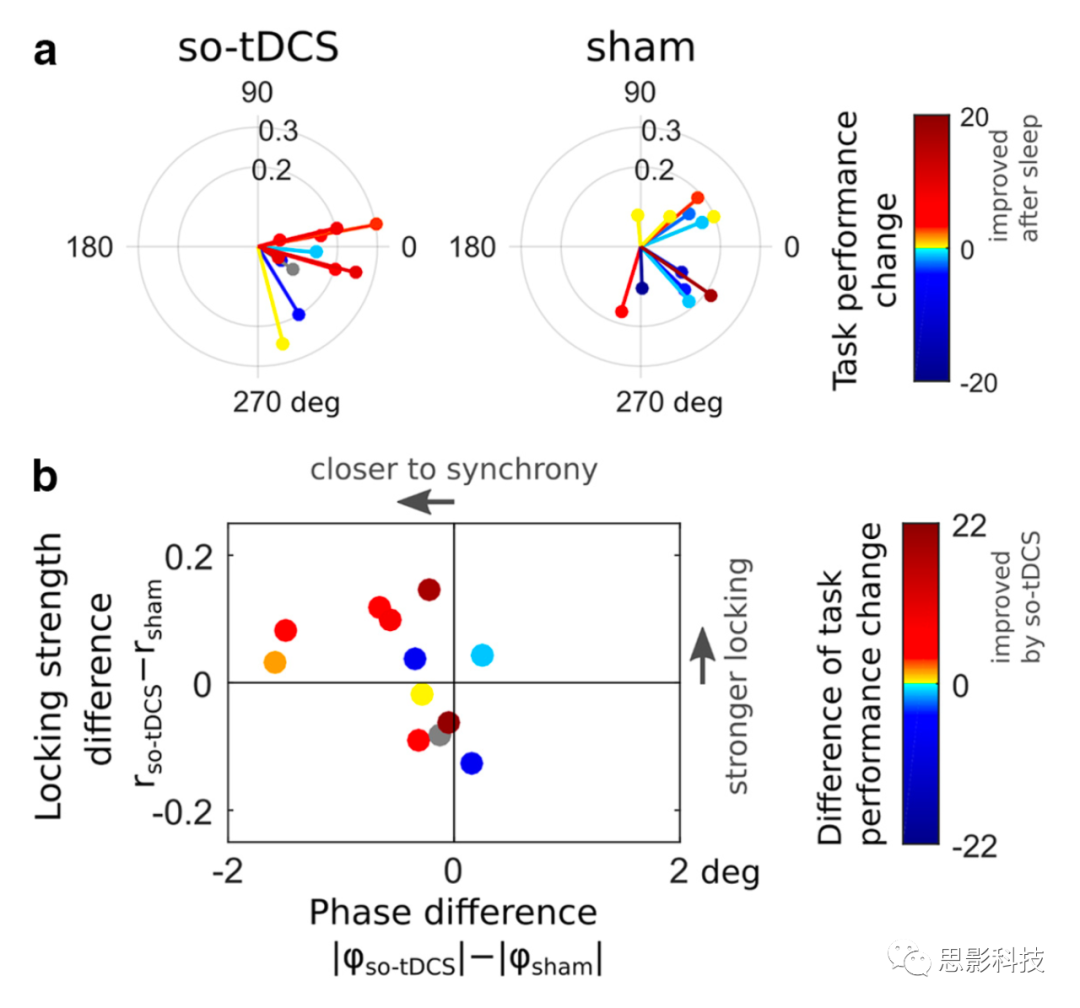
ͼ5 SO����ٷĴ�����λ�����Ϻ��Ӿ������������֮��Ĺ�ϵ��
a.�����߸������붥ҶROI������ƽ��SIֵ��
b.�����߲�ͬ�����µ�SI�뾶����(��������ǿ�ȵı仯)�����SI�ǵIJ��졣
���Ǵ̼������PAC�仯���Ӿ�ʶ�����ܵĹ�ϵ�����ǵ���λ��(SI�Ƕ�)������ǿ��(SI�뾶)��so-tDCS�����ļ������ܸ�������������SO����ٷĴ�������֮���ǿ��ͬ��(SI��λ)(ͼ5b)��֮ǰ�Ѿ����֣�SO������(�¼������½���֮��)�Ŀ��ٷĴ���������(��ͷ)���������أ���ˣ���������Ӧ����һ���dz����ƵĴ�ʩ(���ٷĴ������ʷ�ֵ������SO�����ε�ʱ��)�����Ƿ��֣����ٷĴ������ʷ�ֵ����ʱ��(��SO�����εľ���ʱ���)��̼�������Ӿ�������ֱ仯û�������(�������ڣ�����Pearson 's r����ֵС��0.27������p >0.4)��
�ܽ���һ���֣��̼�������Ӿ�����ĸ���������SO��Ĵ������ʵı仯û������������ԣ�������SO�Ϳ��ٷĴ������ʲ���֮���ͬ������ǿ�йء�
4.����
����Խ��Խ���֤�ݱ���MCI����˯���жϺͼ������֮�������ϵ�������о����ض��ļ������˯�����������ر���SO�����������ٷĴ�������Щ��֮�����������Ƿ������MCI���߰���С˯ʱͨ��so-tDCS��ǿ�����⣬���Ǵ���ͨ���̼��ٽ��û�������Ӿ����乮�̡����ǵ�һ����MCI�����з��֣�so-tDCS��������SO�ͷĴ������ʡ���Ҫ���ǣ�����֤����so-tDCSͨ�����·�ʽ������Դ��SO�ͷĴ��������ǿ���Ĵ���������ȥ����SO�����������Ŵ���SO�Ϳ��ٷĴ��������ź�֮���ͬ���Ը�ǿ�����⣬���Ƿ����Ӿ��������ܵ����������SO�Ϳ��ٷĴ�������֮���ǿ��ͬ�����йء�
���о���һЩ������ֵ��ע�⣺���ȣ���ٴ̼������ȣ���������˯��о������롣������֮ǰ���о��У��ڽ�������������Ҳ���������ֶ���˯��Ӱ�죬��֮ǰû��������˯����Ӧ��so-tDCS���о����������Ӱ�졣���Dz����ų�����˯�����֮ǰ������so-tDCS���µ�SO����ӳ������ڣ��������Ƿdz������ܵġ�Ȼ������ע�⣬���Ƕ���˯������ͳ�ƿ��ƣ����ų���Լ�����ֵ�DZ��Ӱ�졣��Σ�Ϊ�˷���so-tDCS��SO�ͷĴ���֮����ϵ�ֱ��Ӱ�죬�����о���Ƶ�ԭ����ʹ�������൱����EEG����(������1�����̼����)�����⣬����SO���Ĵ�������ʱ��Э���ƺ��Ǽ��乮��������ģ�������������ѧ����(��BDNF�ͷ�)����Ҳ�й��ף����ܲ���so-tDCS�ĵ��ڡ������о�����һ����������������̫С������ܽ�����EEG�ͼ���ЧӦ֮��ȱ����������Ե�ԭ��
�ܽԽ��Խ���֤�ݱ�����˯���ж��ڰ����ĺ�Ĭ��(AD)�ķ�չ����������������Ҫ�����ã�˯���ƺ������ּ������Ƶİе㣬�ر����ڶԿ�����˥�˷��档�����о�֤���������֪�ϰ�(MCI)������˯���ڼ�������Դ̼���DZ�������ƽ������Ƕ�����Ƶ����⡣�����״��ṩ��֤�ݣ�֤�������̼�����ǿ�����������֮��Ŀ�Ƶ��Ϲ��ܣ�������MCI���ߵ��Ӿ����乮�̡�
����ԭ�ļ��������������˼Ӱ�Ƽ��ţ�siyingyxf��18983979082��ȡ,���˼Ӱ�γ̼��������ȤҲ�ɼӴ��ź���ѯ����˼Ӱ�ṩ����������ط�������ҪҲ�����Ӵ��ź���Ⱥ��ԭ��Ҳ����Ⱥ�����������ǵĽ���������о��а����������ת��֧���Լ����½ǵ��һ���ڿ����Ƕ�˼Ӱ�Ƽ���֧�֣���л��

��ɨ����߳���ѡ��ʶ���ע˼Ӱ�dz���лת��֧�����Ƽ�
��ӭ���˼Ӱ�����ݴ���ҵ�γ̽��ܡ�����ֱ�ӵ���������ּ������˼Ӱ�Ƽ����еĿγ̣���ӭ�����ź�siyingyxf��18983979082������ѯ�����пγ̾����ű��������������ǻ��һʱ����ϵ���������ѱ���ѧԱ�����
�Ե缰���⡢�۶���
�Ͼ���
�ڶ�ʮ�����Ե����ݴ������Űࣨ�Ͼ���4.23-27��
������
�ڶ�ʮ����Ե����ݴ������Ű�(������5.16-20��
�Ϻ���
�������Ե����ѧϰ���ݴ����ࣨMatlab�棬�Ϻ���5.15-20��
����ʮ�����Ե����ݴ����м���(�Ϻ���5.28-6.1��
���죺
�ڶ�ʮ����������Թ������ݴ����ࣨ���죬5.24-29��
�˴ţ�
������
����ʮ�Ž�Ź�����Ӱ������ࣨ������4.14-19��
�ڶ�ʮ�Ž�Ź������������ݴ����ࣨ������4.22-27��
�ڶ�ʮ���DTI�ࣨ������5.6-11��
��ʮһ��Ź���ASL�ࣨ������5.12-15��
�ڶ�ʮ������Ӱ�����ѧϰ�ࣨ������5.28-6.2��
�ڶ�ʮ��Ź�����Ӱ��ṹ�ࣨ������6.4-9��
���죺
��ʮ�Ž�Ź�����Ӱ��ṹ�ࣨ���죬4.13-18��
����ʮ��Ź�����Ӱ������ࣨ����,4.21-26��
�����Ӱ����ѧ�ࣨ���죬5.19-23��
�Ϻ���
��ʮ��������̬���ܴŹ������ݴ����ࣨ�Ϻ���4.21-25��
����ʮһ��Ź�����Ӱ�������(�Ϻ���5.7-12��
���£��ڶ�ʮ�˽�Ź������������ݴ����ࣨ�Ϻ���5.22-27��
���£����Ľ�Ӱ����ѧ�ࣨ�Ϻ���6.2-6��
���ݴ���ҵ����ܣ�
˼Ӱ�Ƽ����ܴŹ���(fMRI)���ݴ���ҵ��
˼Ӱ�Ƽ���ɢ��Ȩ����DWI/dMRI�����ݴ���
˼Ӱ�Ƽ��Խṹ�Ź����������ݴ���ҵ����T1)
˼Ӱ�Ƽ������ද���С����Ӱ�����ݴ���ҵ��
˼Ӱ���ݴ���ҵ������ASL���ݴ���
˼Ӱ�Ƽ���Ӱ�����ѧϰ���ݴ���ҵ�����
˼Ӱ�Ƽ������Ⱥ����ҵ��
˼Ӱ�Ƽ�EEG/ERP���ݴ���ҵ��
˼Ӱ�Ƽ��������Թ������ݴ�������
˼Ӱ�Ƽ��Ե����ѧϰ���ݴ���ҵ��
˼Ӱ���ݴ������������Դ�ͼ��MEG�����ݴ���
˼Ӱ�Ƽ��۶����ݴ�������
��Ƹ����Ʒ��
˼Ӱ�Ƽ���Ƹ���ݴ�������ʦ ���Ϻ����������Ͼ���
BIOSEMI�Ե�ϵͳ����
Ŀ��ʽ���ܴŹ���̼�ϵͳ����