���ȥ����ͼ����йص��Իģʽ���Է��طţ�replay�����Ի��һ������������һЩ�����ھ�Ϣ״̬�µ���ɼ���Ҳ���������������ǹ���Ĭ��ģʽ���磨default mode network, DMN����������������Ϊ������������ܽ�֯�����Ҿ���DZ�ڵĹ����ص��������ڴ����������ļ���ϵͳģ�͡���������DMN���طŴ����Ļ��������鵼�˺�������Ƥ��֮�������ã��Ӷ������µļ��䡣DMN�����Զ��������طż�����֧�־ɼ�����ؼ�����߸�������������������Ϊ���ɶ��ݵ�Ƥ�㼤���շ��ĺ���������Ƥ��ij��̹����ǴӶ��¼��ļ�֪�������������������������¼��IJ㼶�����Ĺؼ����������ķ�����Nature Reviews of Neuroscience��־��(�������ź�siyingyxf��18983979082��ȡԭ�ģ���˼Ӱ�ṩ����������ط�������ҪҲ�����Ӵ��ź���Ⱥ��ԭ��Ҳ����Ⱥ���)��
˼Ӱ���������ڹ������䣬DMN��ص����½�����ɽ���Ķ�������������ֱ�ӵ����������������ź�siyingyxf��18983979082��ȡԭ�ļ�������ϣ������䣺
˯���ڼ����ຣ�������������Ĵ����Ͳ��Ƶķֲ�Ƕ��
���ຣ�����л�����Theta-gamma������֧���µ��龰������γ�
��������е�Theta��
Current Biology���������˻��ߵ�˯������
Science�������Ⲩ������SWRs���������Ӿ��龰�����й�
Science�����˺�Ļָ������������Ƶ�����
���������б���״̬������
������˯�ߣ�˯����֪��������ѵ�ͷ�Ա�ø���
ֱ����̼��Դ��Լ����·������ЧӦ
����̼��������Ͳ���˥�Ϲ������龰�����Ӱ��
CELL REPORTS�������Ľ����Թ������伤��
��������Ե��о�������Ϣ���ڸ�֪�ͼ������ܵ��������෴��
���������ڼ���������������Ƥ�㽻���Ķ�̬����
CELL REPORTS�������Ľ����Թ������伤��
theta�������Ѻ�˯�߽εļ����ټ���
����̼��ٽ�˯�����书�������ǿ�����֪�ϰ����ߵļ��乮��
TDCS�̼�ǿ�ȶԽ��������߹��������Ӱ��
֧�ֹ�������Ķ�ģ̬���綯��ѧ
����ѵ��������·�����ǿ�� �������˿־������ȡ�Ļ���
���ͦ����ڹ���������������õ����֤��
�����������֪��ѧ
Neuron���������Ц���ѡ���Լд���������������������
ͨ�����ļд������������
�Ĵ����ڶ�������Ĺ���������Ҫ����
Science: ��������Ƥ��:��ҶƤ��ļ���ӡ��
��˯��ѧ��˯���н��������ʻ�ѧϰ��ȻҪ��������������
�ռ乤������Ĺؼ����ƣ�ǰ��ҶƤ���Theta��High Gammaͬ��
������ѧϰ������Ӱ��
������˯�ߣ�˯����֪��������ѵ�ͷ�Ա�ø���
����̬fMRI�о���TMS�̼�ѡ���Ըı��龰��������Ĺ���
�Ӿ���������ָ������Ϊ�е�ͬʱ�Ӿ����˶�ѡ��
���Ŵ̼�ǰ��Ƥ����ǿ����־���������
�����˺�������������ģʽȱ�ݵIJ�ͬģʽ���Խṹ����
PNAS:��ʱ����˶����ƺ������书��-����̬PPI�о�
alpha�����ӳ�ռ乤���������
���Զ�̬����״̬����Ϊ����Ĺ�������Ļ���
DMN:
��֪�ӹ���ص�Ĭ�������Ա�֢�о��е�Ĭ������
Ĭ�����磺���µĽ��ʡ������о������о���չ�����е��¹۵�
DMN:���Ե�Ĭ������
���Ա��Է�����ס��뾲Ϣ״̬��ȣ����֪����Ϊ��صĻ�ı�����������ĵij̶��Ƿdz����ġ����һЩ���ۼҽ�������Ϊһ����յġ�������ϵͳ���о��̼�ֻ�����ϵͳ�ĵ������ض����Ǿ������ء���ˣ��ܴ�̶������Է�������˴̼������Ľ����Ȼ���������Է������ϸ�ṹ������Ȼ��Щ���ء�
��������������Ҫ��ʵ�鷢��ͻ���˴������ڶ���ѧ����Ҫ�ԡ�����Ⱥ�ͱ�������Ϸ����˼����طţ�memory replay����������ǰ�Ļ�����з����ļ���ģʽ���Է��ؼ����������Զ���ѧˮƽ�������˾�Ϣ̬������Ĵ��ڣ����ڲ�ִ�л�����ڼ�һЩ������Ի�������������һ����Ϣ̬�����磬DMN�������������ע�⣬��Ϊ��������״̬�ͼ�������йء�
���������Դ����ѧ�зdz���ͬ����������������ѧ����Ҫ�о������ද���������֪��������Ƕ�������������Ϊ�طź�DMN֮�������Ҫ��ϵ����������һ���۾��н���ѧ�����ܺ�������֤��֧�֡����������������ʱ�ճ߶Ⱥͷ�����������ȫ��ͬ�ģ������������еĵ�������Ҳ��ȥ��Զ���ӹ��ܵĽǶ�������DMN��Ҫ������ȥ�ľ��������Ҳ��յ�������йء���һ���棬�����طţ�����Ϊ֧���˼����ϵͳ���̣��������������е��µĽ��ʺ����ӽṹ���Ա㳤�ڴ洢���ʹ���ؾ�������ȡ��һ��֪ʶ���γɣ�������䣩��Ȼ��������ķ���ѧ��չ�õ��Ľ������������������֮�������ϵ���������ǵĹ��ܡ��漰�Ľ�������Ͷ���ѧ����������ԡ����ǵ���һ֤�ݣ�������Ϊ�طź�DMN�ʵ���Ͽ�����ͬһ���䴦��ϵͳ��һ���֣�ֻ�Ǵ����˲�ͬ�Ƕȡ�
�طź�DMN֮��Ĺ�������Ҫ�������еļ���ϵͳ����ģ�͡�����ѧϰϵͳ������complementary learningsystems theory����Ҳ����Ϊ����ϵͳ���̵ġ�������ģ�ͣ��������ӵ������ר�ŵ�ѧϰ�ͼ���ϵͳ��һ���ں����壨���ٻ�ȡ����Ϣ������һ������Ƥ�㣨�洢���������µ���Ϣͨ��һЩ�д�������ݶ��½��Ż��㷨������ͼʽ֪ʶ�⡣��һ���۱�����������Ϊÿһ�����صľ�������һ�����ص�������룬��ơ������������������������һϵ�й㷺�ֲ��ĵͼ�Ƥ�㼯Ⱥ����ЩƤ�㼯Ⱥ�����������������ԣ���ʵ�����Ϻͼ��������ݡ�������ģ�ͣ��طŵ�������ʹ��������ϵͳ֮����Խ��н������Ӷ������ؽ�����Ϣ���뵽��Ƥ�ʵĴ洢�У���ֹ��ѧϰ�²���ʱ���������Ը��ţ�ͻ�����ӱ�����Ϣ��ûʱ�����洢��Ϣ����ȫ��ʧ����Ϊ��ʵ����㣬����Ϣ���طţ�Դ�Ժ������;���Ϣ���طţ�Դ����Ƥ�ʣ����뽻�����С�
���Ǵ˴������������۵ĸĽ��棬��������ļ���ϵͳ����cascaded memory systems, CMS��ģ�ͣ����ǽ���������Ĵ������طŵķ�����չ��һ�������������DMN�ͺ��������硣������Ϊ������������������ط�ͨ��һϵ�д������缶�����������ȫ�Բ���ġ����Էḻ�ĶԾ���ı���������γ���һ���ֲ�����ϵͳ�������ϵͳ�У�����ÿ������Ĵ����������Է�������ض���ѧ���ȿ��������洢�ڽϵͼ��ڵ��еı�����ͨ�����϶��µĹ��̣����ֿ���ͨ���������������������¼��ͨ�����¶��ϵĹ��̣�������������ȫ�ּ��������ǽ�DMN��Ϊ��������طŵ����ģ���������Ϊ�������������Ե�ͨ�á�����˫���ͨ·��CMS������ļ���ϵͳ��ģ�͵���һ����ӱ֮��������ǿ����Ƥ�ʶ���ѧ����һ�������������ٽ��ԣ�criticality������֪ʶ��1��������Ϊ��νṹģ���п�Խ�����ε���Ϣ���塣���������������CMSģ�Ϳ��Խ���һЩ�����ʵ��۲��������������ڴ��Ի�ͼ�����µĿɱ�����Ԥ�⡣
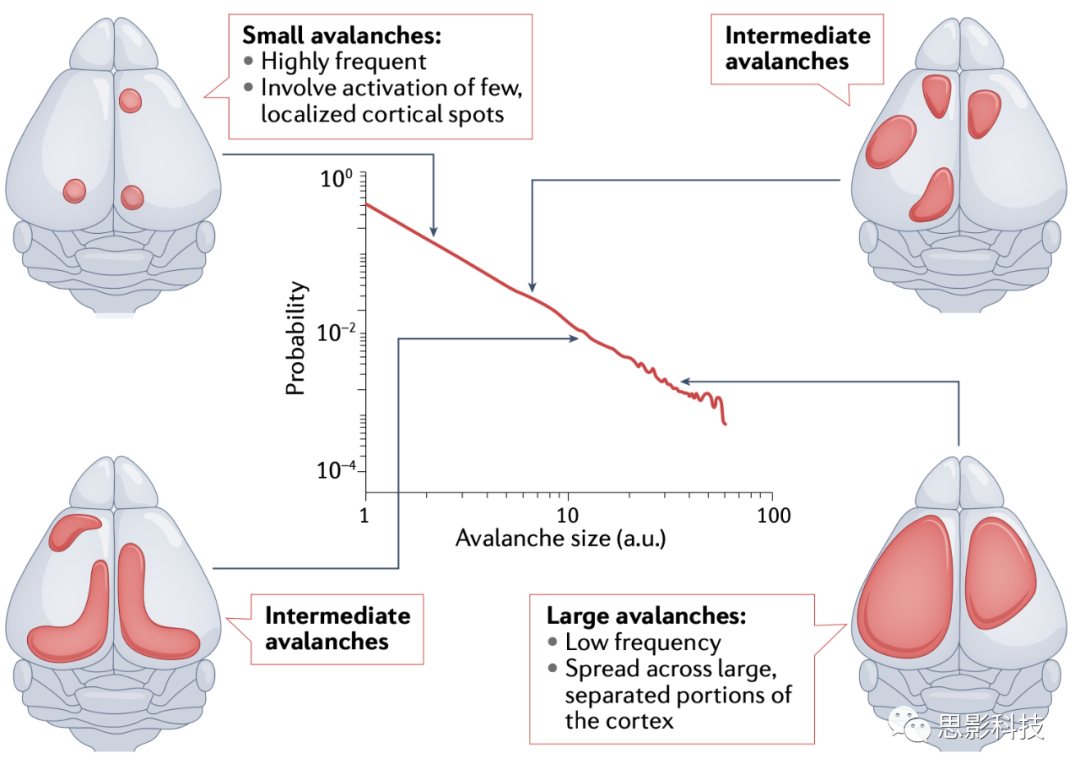
| ֪ʶ��1 Ƥ�㶯��ѧ��һ�����ۣ��ٽ��Ե�����
�ٽ��Եĸ������������ѧ����������ѧ�����ٽ��Եķ�չ��Ϊ��ʶ���ɴ���ʵ����ɵ�ϵͳ�ļ�����Ϊ���ٽ���������һ�����������֮�����ȫ���������һ����֮��ﵽƽ���״̬��������ٽ���Ϲ�����ϵͳ���ص���ϵͳ���֮������ų����������ԣ���ʹֻ�зdz��ٻ�û�г��̵����ӡ�������ѧ�ĽǶ��������ٽ�ϵͳ�������ǻ��ʱ�ռ�������Ϊѩ�����䷢�����������С�����ʱ�仺���½������ɷֲ����μ�����ͼ���ܺõ����������ֱ仯������ζ���������¼��У������¼�ռ�������ľ��֡���ˣ�ϵͳ��ʱ�䶯��ѧ���ձ���Щ�����¼���������
���ֶ�ϵͳ����ѧ����������ѧ�еõ��˹㷺�Ĺ�ע����Ϊ���������������������ͨ��������ʽ�Ŀ����Խӽ��ٽ�״̬���Լ����۵�ʵ��֤�ݱ������ڸ������͵�Ƥ�ʼ�¼�Ļͳ�������У������ٽ������������ʵ����ɱ�ȣ������ٽ�ϵͳ����ȿ��������ڽ��������������淢�֣������в���travelling waves���Ĵ�����Դ��DMN�����⣬�ﵽ�ٽ�״̬����Ϊ�������������Ŀ�꣬��Ϊ�����������Ϊ������������������ԣ�������ⲿ�̼�����Ӧ����Ϣ�������Ϣ�洢��
Ȼ�������������Ƥ����Ⱥ�ﵽ�ٽ�״̬�ij̶ȱ����������������о���������Ⱥ��ñ�����Ϊ���������죨reverberating����״̬���Ե����ٽ�ֵ����ˣ������п������ٽ��ԵĻ���Ӱ��֮��ȡ��ƽ�⣬ͬʱ���ⲻ̫�����Ӱ�죬���������ԺͿɿ��Խ��͡�Խ��Խ���ʵ��֤�ݱ��������ֽ��ٽ綯��ѧ�����������DZ�ھ�̬�ṹ֮������Ź�ϵ���ر��ǣ�DMN������ά�ֽ��ٽ����綯��ѧ��ͨ�����������ٽ�ѩ���¼���Ч�������淢������Ҫ���á���Ȼ�����������������ͽ��ٽ�״̬֮��Ĺ�ϵ����Ϊ�������ʱ�겨����η�ӳ�ģ����У���Ŧ������DMN�����ɲ�������Ҫ�Ե�Ƶ�����е����綯̬��
|
�����ط�
�����ط������ݶ���˯���ڼ�ĺ������б����緢�ֺ���ϸ��������������λ��ϸ���������ڶ����Ļ����У�ÿ��ϸ�����в�ͬƫ�õķŵ�λ�ã���Ҳ��������ʱ�о���������Է����ڲ�״̬���罱������������������ж��ƻ����ĵ������������˵�ǰ���������е����ģʽ��˯���ڼ亣��λ��ϸ����ͬʱ�Ͱ�ʱ��˳�����ط��ں�����Ԫ�ϵļ�¼�б��״���������������˸�����չ���е��طţ����������״̬�켣����ĺ���ϸ����ͼ1a����
��Щ���㷺�Ļط����е�����������Ҫʹ��λ�ý��룬����һ��ͳ�Ƴ���ͨ���ó�����ԴӼ�¼����Ԫ��Ļ����ȡ����ĵ�ǰλ�á�ͨ�����Ӷ������Թ��б���ʱ��¼����Ԫ��л�õĽ�����Ӧ����ͬһ������˯���ڼ�ĺ�����¼�����������˯���ڼ䷢�����Է����ѹ����ʱ��߶������������Թ��е����Ҳ����˵��˯���еĺ�����Զ�������ʱ��Խ�Թ�ʱ��������ͬ˳��������λ��ϸ���ļ������С��뺣�������ȥ������������뷨һ�£�������������ͷ������Ҳ��֤������˯���طű���ġ�
�ں������ط����ȷ�����SWRs(�Ⲩ����)�ڼ�������������Ԫ��30�C100���������������ɵĶ������з���Ĵ����ϵ������¼�����ӳ�����ǵ���ͻ���룬���ϵ����˸�Ƶ��~200Hz����ϸ�������ƣ�����ӳ�����������SWRsÿ�뷢�����Σ��ɺ���CA3�������˷��Է����������������зḻ�Ļ�ʽ���ӣ������ҿ��ܱ��Է���Ƥ���շ���SWR�ڼ�۲�طŵĸ������ӣ�ͼ1a��֧�������Ĺ۵㣬�������������ܻᴥ����ͻ�������б���ļ���켣�ļ�����
��ǰ�����˯���ڼ亣���طŵ������У���������������ѧϰ����Ϣ�洢ģ���������ļ����������ƥ�䡣�����֤�ݱ���������Ļ��ʺ���֯������ʱ��ı仯���仯����Щ���֣��ټ�������ģ�ͣ�����ϵͳ�������۵��γɺ��طű���Ϊ�Ǻ�������Ƥ�ʼ���ϵͳ֮����Ϣ������DZ�ڻ��ơ���ʵ�ϣ�SWR���طŻ�����Ʊ�֤���ᵼ�¼���ȱ�ݣ�֪ʶ��2��������ԭʼ�������ʽ�У�ϵͳ������������˼��乮�̹����м���Ӻ�������Ƥ�ʵ�����ת�ƣ�����Խ���һЩ���֣����纣�����˺����˳������������Ȼ���������γɵ��������۲�û�м�����䱾������ϸ�������κ�ʱ����һ�ֶ����ķ�ʽ�洢�ں������У�����������ɺ������ģʽ�㷺�ֲ���Ƥ�����Ա���֮�����ϵ��ɣ�������ҪƤ��-Ƥ�����ӵij��죬�Ӷ����ͺ�����������Ҫ�ԡ������ط���ΪƤ�ʿ����Ե�һ���źű���Ϊ�����ֳ������������
��SWR���ط�Ҳ�����������ںͳ���������Ĵ�������ʱ���ط�����ȡ���˽�һ�����ش��չ������״̬�µ�SWR�Ͱ�����ط�����������ڼ�ż��������ͨ���ǵ�������Ŀ�������ѡ��������ͣ���������Ϊʱ���طſ�����˯��֮����У����ҿ���������ǰ���������������������Ͻ��У���һ���ֵ����طž��и���Ĺ��ܡ��������ڼ��乮���е������⣬������Ϊ�������ڼ�ĺ����طſ��������ڻ�����䣬Ϊ����ģ�⡢�ƻ��������ṩ������֪ʶ��2������SWR�ڼ䣬���ຣ����Ԫ��ͬ���ŵ��ṩ��һ��ǿ���Эͬ�źţ���������������������������Ϣ�������������ȡ��Ϣ����ʵ�ϣ���Ҷǰ����Ƥ�ʺ���Ҷ���Ӿ�������Ƥ��������Ҳ�����뺣��SWRsһ����ֳ�����Ļ���طţ�ͼ1b,c���������ڣ���Ƥ��Ҳ�б����ᷢ��SWRs��ͼ1d��������Ҳ��ط��йء��ܵ���˵����Щ���ݱ����طŶԼ�����̵Ĺ��ײ��������ں����������漰��ȫ�ԡ�
���ݶ����о��Ĵ������ֽ�SWR���ط�����������ϵ���������������з��������Ƶ������������У�SWRҲ�����ں������ڲ��Ҷ���ڽ������������ֳ�����ء������������н��������Ե���Ԫ��¼ͨ�������У�������ܴŹ������fMRI�����ݵĶ��������������֤���������طţ�����������˯�ߺ�����״̬�µĻ�����طš���������ط��о�����֤ʵ�����ݶ��������֣�Ҳ������֧����Ƥ���ط��źſ��Զ����ں��������Ĺ۵㡣���˷�����Ƥ���ڵļ�����̲������µĺ��̵ļ��佻���������һ���о��������Ƥ��SWR����ͬ����Ƥ���������������طţ������Զ�̼�������з�����Ҫ���á��۲쵽SWR������������Ƥ�ʣ�˯�ߺ�����ʱ��������������Ƥ��λ��ͬʱ��������Ϊ�������Ƥ������ֶ����ط��ṩ��֤�ݡ����⣬�ɹ��ļ����ӳٻ��䲻���뺣��-Ƥ�ʵ�SWRs��Эͬ�����йأ������Ƥ��-Ƥ��SWRs��Эͬ�йء�
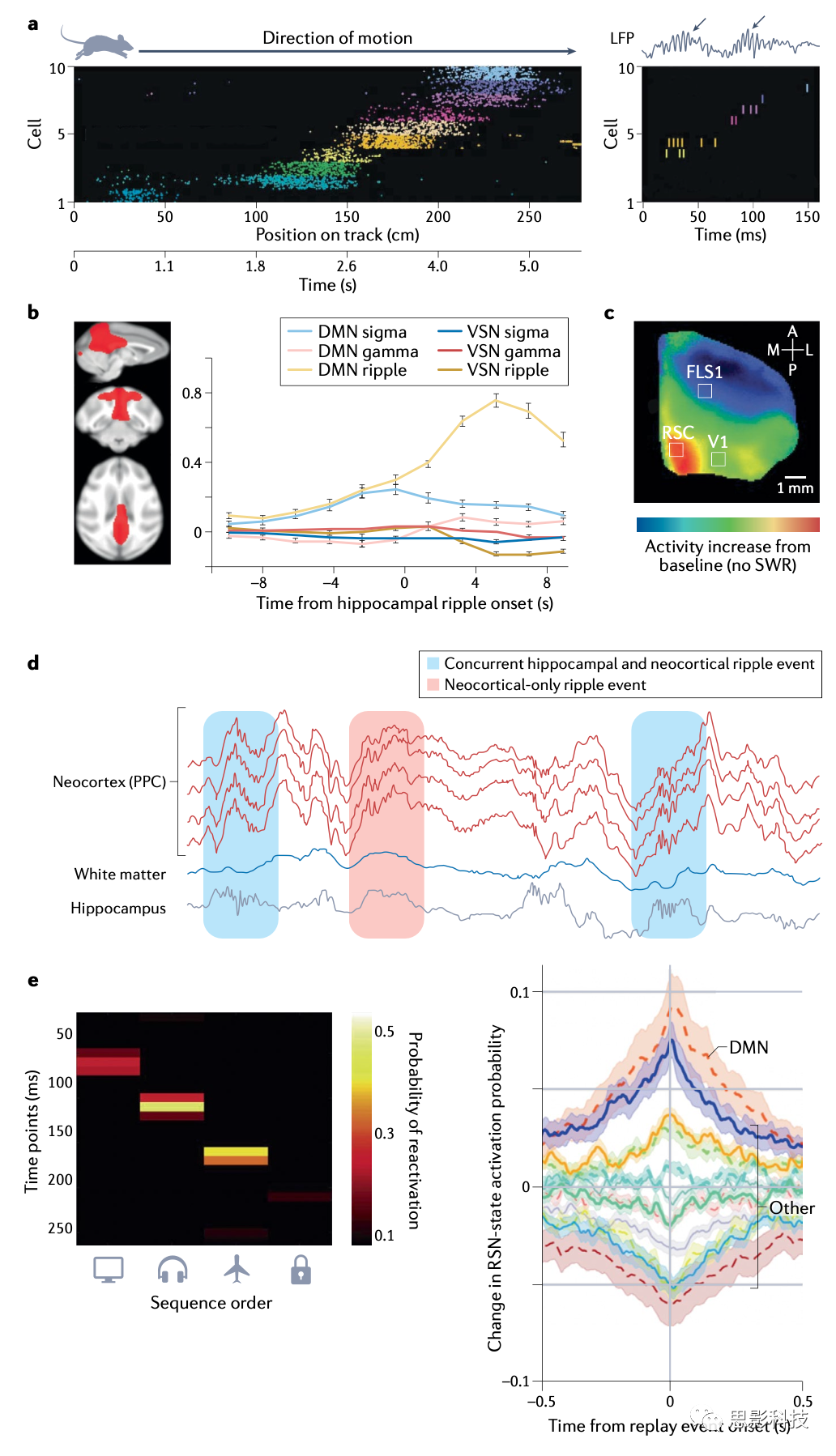
ͼ1 ��Ϊһ��ȫ������ļ����ؼ���
a. ���ݶ��ﺣ���е��������ݼ����ڼ���������SWR�����ڼ�ķ�����������״̬�£����������Ź켣�ƶ�ʱ�����������е�ϸ����˳�����С�ÿ���̶ȴ���һ��������λ������ɫ�ʹ�ֱλ��ָʾ��¼����ϸ����������˯���У�ͬһ���е�ϸ����ʱ��ѹ���ķ�ʽ���¼����ͼ���������ط�ͨ���뵥����SWR��Ӻ����ֲ�����λ��LFP����¼�Ķ��ݡ�������SWR���У���ɫ�켣�ͼ�ͷ��һ������
b. ����SWR��������Ƥ�ʻ��ͬʱ���ӣ�������DMN���������ʾ�˺�Ӻ�DMN�ķ�Χ���Ժ�ɫ��ʾ�����Ծ�Ϣ״̬��������Ϊ�������Ҳ���ʾ���ں���sigma��8-22 Hz���ڼ��������DMN���������˶����磨VSN����һ����Ƥ�ʾ�Ϣ״̬���磩�ռ���fMRI��¼��gamma��25�C75 Hz�����Ʋ���80�C180Hz���������SWR��DMN����Ļ��ǿ��һ�¡�
c. С����Ƥ�ʵĻ��ʹ�õ�ѹ����Ⱦ�ϲ����������ຣ��SWR��ֵʱ���ƽ��ֵ����Ƥ�ʣ�retrosplenial cortex, RSC����DMN��һ���֣���ʾ����SWR�������̶߳ȣ�����̵���ʼDZ���ڣ�δ��ʾ����
d. ��Ƥ��Ҳ����������������500�����˯��¼�Ĺ켣��������ҶƤ�ʣ�PPC����SWR�����뺣��SWRͬʱ��������Ҳ������������ɫ�켣��ʾ�˰������Ŀ��ƻ��Ԥ�Ʋ������������
e. ������Ƥ�ʵ����лط���DMN�ļ�����һ�¡������߱���ѧϰһϵ���Ӿ���Ŀ����������Ϣ�ڼ䣬ͨ��ʹ�ö���Դ�ͼ�������������Ƥ�ʼ�¼�Ļ�����Լ���Ŀ���е��طţ���ࣩ���طű���������Ϣ̬�������еĻ��һ�£�DMN����ɫ����ʾ����ǿ�IJ����������Ҳࣩ��������¼����ʾ������Ϣ̬�����硣
��������Ӱ�������������ݴ������Թ��ܣ������������ģ̬��Ӱ�����ݴ�������Ȥ�������˼Ӱ�������ӣ�ֱ�ӵ���������������лת��֧�֡�(�������ź�siyingyxf��18983979082��ѯ)��
�Ͼ���
����ʮ����Ź������������ݴ����ࣨ�Ͼ���10.16-21��
�ڶ�ʮ����Ź�����Ӱ��ṹ�ࣨ�Ͼ���10.24-29��
����ʮһ��Ź�����Ӱ������ࣨ�Ͼ���11.12-17��
�Ϻ���
�ڶ�ʮ�Ľ���Ӱ�����ѧϰ�ࣨ�Ϻ���10.9-14��
����ʮһ��Ź������������ݴ����ࣨ�Ϻ���10.28-11.2��
����ʮ�Ž�Ź�����Ӱ������ࣨ�Ϻ���11.4-9��
���죺
�ھŽ����������ݴ�����߰ࣨ���죬10.13-18��
����ʮ��Ź�����Ӱ������ࣨ���죬10.22-27��
�ڶ�ʮ�˽���ɢ�������ݴ����ࣨ���죬11.5-10��
��������ɢ�Ź��������߰ࣨ���죬11.17-22��
�ڶ�ʮ����Ź�����Ӱ��ṹ�ࣨ���죬11.27-12.2��
������
��ʮ�����������ݴ�����߰ࣨ������10.20-25��
��ʮһ��Ź���ASL������������ǣ����ݴ����ࣨ������11.3-6��
����ʮ����Ź�����Ӱ������ࣨ������11.9-14��
������Ӱ����ѧ�ࣨ������11.25-30��
�����R����ͳ�ưࣨ������11.16-20��
���ݴ���ҵ����ܣ�
˼Ӱ�Ƽ����ܴŹ���(fMRI)���ݴ���ҵ��
˼Ӱ�Ƽ���ɢ��Ȩ����DWI/dMRI�����ݴ���
˼Ӱ�Ƽ��Խṹ�Ź����������ݴ���ҵ����T1)
˼Ӱ�Ƽ����������У�QSM)���ݴ���ҵ��
˼Ӱ�Ƽ������ද���С����Ӱ�����ݴ���ҵ��
˼Ӱ�Ƽ��鳤�ද��fMRI����ҵ��
˼Ӱ���ݴ���ҵ������ASL���ݴ���
˼Ӱ�Ƽ���Ӱ�����ѧϰ���ݴ���ҵ�����
˼Ӱ�Ƽ������Ⱥ����ҵ��
��Ƹ����Ʒ��
˼Ӱ�Ƽ���Ƹ���ݴ�������ʦ ���Ϻ����������Ͼ������죩
BIOSEMI�Ե�ϵͳ����
Ŀ��ʽ���ܴŹ���̼�ϵͳ����
DMN
�ֲ�ʽ���������е�ʱ����ػģʽ����Ϊ�����˹������硣������Դ̼�ͬʱ������Ӧ�������Ĵ�������֮�����ػ�⣬�ھ�Ϣ״̬�£�����û�������̼�������£������۲쵽�Է�����������ԡ������о��Ѿ�ȷ���˼�����ʾ���������Է����ϵͳ��ͳ��Ϊ��Ϣ״̬���磩�����������˶����Ӿ���������ע����ϵͳ����������Ϣ״̬��һ�����ھ�Ϣ״̬�£�DMN������Ҳ��Ѫ��ˮƽ������BOLD���źŵ������룩ʱ��߶�����ʾ��һ�µĻ��Ȼ������������Ϣ״̬������ȣ�DMN�Ƕ��صģ���Ϊ��������»����ʱ�����ļ���ˮƽ�ή�ͣ������ע��������ʷ���أ���
����������ص㣨����ʹ�������ӷ���ϲ�ɨ�裨PET���ͺ����Ĺ��ܴŹ������fMRI������DMN�����������ͱ����ڲ�ǰ��ҶƤ�ʡ���۴���Ƥ�ʡ�ШǰҶ�ͲඥҶƤ�ʡ��ڲ��Ҷ�ͺ���Ҳ������DMN�С����磬DMN��Ƥ�������Է���뺣�����Է����ء�DMN����֮��Ĺ�����ؽṹ��˯���ڼ䲿�ֱ����������ȶ��Ա�����DMN������ij�̶ֳ��ϣ�״̬����������֪�Ľ�������һ�¡��������½��е�fMRIʵ��Ϊ�������鶯���д������Ƶ�DMN�ṩ��֤�ݣ�����С����è�ͺ��ӡ�
����DMN�Ĺ�����ȱ����ȷ�Ĺ�ʶ����֮���Ա����֣�����Ϊһ��ʼ���о������ʾ������û�б������ض�����ʱ��һ���������ӣ�����ζ��DMN��������Ϊ��ӳ����û���ⲿ������ص������������Լ�����뷨��Ȼ�����������������ⲿ��������DMN�е�BOLDʧ����ڱ��ձ���ܵ��ǣ��������ڿ���״̬�´��ڻ״̬����ʵ�ϣ������ڲ�������Ϣ������Ҳ������DMN�ļ������DMN�еĻ��ӳ�����������ڼ����۾��鱨�����ϸ�̶ȡ����磬Ҫ�����������˹�ȥ�������о����֣������龰�������������Ĵ���������DMN�������кܴ���ص�����Ȥ���ǣ������о������������ȥ�Ĵ�����������������δ����������Ҳ������Ϊ�������������Ҫ���δ��Ե�������ǰ���롣ֵ��ע����ǣ�DMN��һ��������������ĸ���������BOLD�źŲ����е�����ԣ���BOLD������ʱ��߶ȣ��뼶�������еģ����ܶ�SWRs�������ڰٺ��뼶�����fMRI���ܲ�����DZ�ڷ�����SWRs�����ʵ�ϣ�ʱ�侫�ȸ��ߵ��Դ�ͼ�о��Ѿ�������DMN���ط��ڼ�Ļ��ͼ1e����˵��������ijЩ����£�DMN�Ҳ����ʵ��Ҳ��ӳ���طŻ��
�������ᵽ��DMN�������ƵĶ�������о�Ҳ�У������ں����м�¼��Ҳ����Ӧ�˶�δ���˶��켣�����Ļ���С����ֻ�Ƿ�Ӧ�˼ƻ�����������м��书�ܣ��������顣��ʵ�ϣ��������ط��ڼ䣬�����ֹ��ܿ��ܷ����ڲ�ͬʱ�䡣
����ģ��ͨ�����зḻ��ϸ�ڣ���˿��Լ�������������������;������Ա�ʾ֮�����ϵ��������������߽������뱻��Ϊ���ں��������ɺʹ洢�ġ���ˣ����ǿ��Լ��躣�������ṩ������DMN�ṹ������-�������ӣ��а�������ڼ���������ص�����������DMN�ڵĺ�����ӳ������֪ʶ����֯������-���������ԣ��������IJ��ֿ���ʵ�ֱ��������ģ�⡣
DMN���������Ļ����йأ��������ij��ڴ洢�йء�����DMN�����������壩�ڵĻ��߹��������볤���龰���䱣�ֳ�����أ�������ڼ�������ء����ֹ�ϵ�Ƿ������ھ��нϸ�DMN���������Եĸ���ļ������¼����������£��д��о������⣬��ͬһ���о��У����ֳ��Ϻõij����龰���䱣�������ĸ���Ҳ���ֳ���ǿ��DMN���֪���ⲿע������Ļ��߲�ƥ�䡣
��֮����Խ��Խ���֤�ݱ�����DMN�����ڱ���״̬�¼�����һ����������йء���Ȼ������Ѫ���ij����ܹ�ͬʱ���������Գ���ʱ��ֱ���Զ���ڵ�����ѧ�ķֱ��ʡ���Ȥ���ǣ����ô��·���������MEG�о�������Խ�ض����������˲ʱ�����������ڱ��״λ���DMN��������ͼ��Ѫ���źŲ��������ʱ��߶���������������������DMN��ǰ���ͺ����ص�������DMN����ѧ�ڿ���ʱ��߶��϶��з�����
����-Ƥ�㽻��
���˴ٽ��طţ�SWR���������Ǻ�������Ƥ��֮������õ���Ҫͨ�����ڷǿ����۶���NREM������˯���ڼ䣬�����������Ԫ�뺣��SWRsһ�µ������˷ŵ硣NREM˯���ڼ��SWRҲ��Ƥ��˯�߷Ĵ����йأ�Ƥ�ʻ�е�9�C20 Hz������乮���йء�����ʱ������SWRҲ��Ƥ�ʼ�������ӻ�����йء�
������NREM˯���ڼ�㷺�Ĵ��Լ�����أ���SWR��ؼ���Ҳ����һ���̶ȵ������ԡ�һ�������ʹ���ľ�Ϣ״̬fMRI�������ѧ���ϵ��о���ʾ�����������ȣ�SWR�ڼ�������Ƥ������ļ����������ӣ���Ƥ���½ṹ����Ȥ���ǣ������Ӿ�Ƥ��Ҳ���٣���������Ҫ���ǣ���ͬһ���ݼ������·����������������ڼ䣬����DMN�뺣��SWRs�����һ�µؼ��ͼ1b��������С��ĵ�ѹ���г��������ṩ�����Ƶ�ͼ�����и��õ�ʱ��ֱ��ʣ�������DMN�������Ǻ�Ƥ�ʣ��ļ�������ӳٺ͵��º���SWRs��ͼ1c�������⣬��������DMN������ֱ�ӵĽ���ѧ��ϵ�������ھ�Ϣ״̬����֮�⣬��DMN�����ȵĹ�����ϵ��ֻ�������˶�����Ҳ�뺣�����л����ԣ�����֧����DMN�뺣����������ϵ�Ľ��ۣ�����SWR��Ϊ��������õ�DZ�ڹؼ����ơ�
�����һ��MEG�о�������Ƥ���طű����������ʮ�����DMN˲ʱ����һ�£�ͼ1e������ˣ����Ǻ����ײ²⣬DMN��������ܴ�����ǰδ�����ֵ��طţ���ΪDMN�о���ͨ��ʹ�õķ�����ʱ��ֱ��ʽϵ͡���ʵ�ϣ������ݶ��������������ѧ�о�������������DMN�ڵ����طŵ�֤�ݡ��ڲ�ǰ��ҶƤ���Ǻ����������ҪƤ������֮һ����������ط�����˯�����״μ��ġ��뺣���ط����ƣ��ڲ�ǰ��ҶƤ���طŵ���ʽ��ʱ��ѹ���ġ�����30�C100����Ķ��ݻ���������ط��������Ϊ��صĻ���С����⣬ǰ��Ҷ���ط�˲�������뺣��SWRsͬʱ��������֧����SWR�����ں�������Ƥ��֮�����Ϣ�����Ĺ۵㡣�ط�Ҳ�������ṹ�б�֤��������Ƣ��Ƥ�ʺͶ�ҶƤ�ʣ�DMN���������֣���
ֵ��ע����ǣ�����������DMN��Ƥ������Ҳ���ֳ��طš������ᵽ��MEG�о���ͼ1e���������й㷺���طţ�����DMN�����������������ͬ���������ݶ����о��У�Ҳ�۲쵽���Ӿ����������˶�Ƥ���Լ��ڲ��Ҷ���������֣�������Ƥ�ʵ��طš���ˣ�DMN����Ϊ����-Ƥ��������ṩ�ӿڣ�ʹ�ط��ܹ������������м�����ʵ���ϣ�Ƥ���ѹ�����ʾ��SWR��صļ����Ƣ��Ƥ����ɢ���������²���Ƥ������ͼ1c��������������Ļ������ǿ���Ԥ��DMN�ṹ�е��طŽ��뺣���ط������أ������о��˶�Ƥ�ʵĻ֮�����ϵ��ȡ����DMN�鵼����һ�������ͨ���Ͽ����ӻ�ʧ������м��顣
| ֪ʶ��2 ���Ѻ�˯�����طŵIJ�ͬ��������
�����طſ��ܷ��������ֽ�Ȼ��ͬ����Ϊ״̬�У����Һܿ����벻ͬ�Ĺ����йء�˯���еĺ����ط������Ĺ�����ϵ��һ��Ȼ�����Ժ��ʱ�����䡣�ⷽ�������֤������һ���о������о��������˴��������ռ�����е�һ���ĺ����طţ����¶��ܸ��ż����ѡ���Լ���ȱʧ������һ�����䱣����á������ں����طž��в�ͬ�Ĺ��ܡ����磬�������ද���У��ڵ�����ͣ�ڼ�����Ѻ����طű�֤������Ԥ����һ��·���������ǰհ�Լƻ����������Ȥ���ǣ����Ѽ����طŲ�����ǰ�ƽ���Ҳ����˵�������¼����������˳�����ҿ��Է���������Ϊ�������طŵ����ַ���켣���Ӷ���ĵ�ǰλ�ÿ�ʼ���ڶ�������λ�ý���������ʵ�ֻ���������乮�̡������طŵ�Ƶ�����Ž�����С�����Ӷ����ӣ����ܻ���ǿ����Ҫ�¼��ļ���ǿ�ȡ�
��Ƥ���طźܿ����ڹ�����Ҳ���Է�Ϊ˯�������ڼ䷢�����¼���Ƥ���ط���˯���乮���е�������δ��ȷ֤����Ȼ������֤�ݱ�����Ƥ�ʻ����طŵ���Ϊ���á����磬�����ݶ�����Թ�����ԣ�����ת���������У�ǰ��ҶƤ���ط����Թ�λ�����У���Щ�����ط��������Թ��켣�������ټ�����ڶ������ŵ�������ʱ�ڣ�������ѡ����Ŀ�����������طŵ�Ƶ��������л����ֳ�����ء����⣬�����ѡ����ȣ������ﴦ�������ѡ���ʱ�������طŸ��п��ܷ������������ﴦ��Ŀ������ʱ�������طŸ��п�����ѭ����ѡ������顣���������Щǰ��ҶƤ���¼������ڼƻ������������ж���������ȥ��ѡ���淢������������DMN������������ǰ�۴��ص������ط�Ҳ���������á�
|
��������ϵͳģ��
�����������������طź�DMN������Ա���Ϊ�Է����ͬ���Ƶı��Ϊ���������ṩ֧�֡���һ���轫�����ǶԼ����Լ��Է�����ط��������õ��������������ҪӰ�졣
������������ϵͳ���乮�����۵Ŀ���ڣ������ط�����������������ϵ������ѧϰϵͳ���۽���һ˼��������һ��������㣨���㣩������ļ���ģ���У����к������ǽϵͲ㣬���������¼����֪��ͳһ��ʾ��ʽ���ʱ�����Լ�����б��롣�ϲ���Ƥ����ѧϰ����֪�IJ�ͬ�ɷֲ�������з��ࡣ�����������ط�ʹ��Ƥ�ʼ���ϵͳ��������ѵ���ܹ�ʹ�µĺ;ɵļ�����ϵ������Ϊ��ʵ����һ�㣬��������Ϊһ��������������¼��IJ�ͬ���Ա�ʾ�ʹ洢�ڲ�ͬ��Ƥ������IJ�ͬ���Ա�����ϵ�������Ӷ�ʹ�����ܹ�ͬʱ���¼��
Ȼ����Ŀǰ�в�����ڽ��ʽṹ�Ϸ�ɢ�ĵĺ����������������ңԶ��Ƥ������֮���յ���Щ����ԡ��ڼ����γɹ����У������������ɹ�ͬԭ����������¶��ϵ�ͬ����������ġ����磬��ʳ�ߵĽӽ����ܻ�ʹ����о��������Ұ����һȺ�������ܵ�ͬ�࣬�Ҳ�Զ����Ұ����һ���ڰ�����в����Ӱ���Լ��������ζ��������Զ������������Э���ķ�ʽ���¼��������Ϊһ������ɵ�Ԥ��ģʽ�С���صļ��ͨ����Զ����ģ����ڱ��������ϵ�ͳ�ƹ��ɣ����磬����ζԤʾΣ�գ�Ҳ�DZ�Ҫ�ģ����ǹ�������ʵ�����������ݡ����˾���Լ������Щ���״̬���ܺ��Ѵﵽ��ά�֣����ͨ���طſ��Լ�ǿ����Ϊ��ֲ�������ȣ���Ƥ�ʵ���������ӳ̶�������ޡ�
����������DMN�ڽ��ʺ����Ͻ������������뺣����������Ĺ�����ϵ�����⣬DMN��ȫ���ط�˲��ͬʱ���������ڵ㱾��Ҳ��ʾ�طš���ʹ�����Ǻ�����Ȥ��DMN��Ϊ�����壨����Ϊ���¼��ز�����Ҫ�����ߣ���֧�ָֻ�����Ƥ���������ķ�DMN����Ƥ������֮������õĹܵ������⣬DMN���������Զ����������²��Ž��ϵ�Ƭ�Ρ��⽫Ϊ����ѧϰϵͳ�����ṩ������ϡ������������Ľ����������������DMN�ڴ��������νṹ�ڶ����е�λ�ã����е������������й�����IJ�ͬ�ӽ��̶ȼ������ϵ���Ϣ���������У�ͼ2������Marr�����������ϵ���һ��νṹ�У�DMNλ�ں���������߽ס��������ı���ͷ�Χ��Ϊ�������ݶ���һ�˵ı�ʾ֮�䡣Ȼ��������Ȼ���ܻش�һ�����⣬������Ƥ������֮��ij������������������ȱ�ѧϰ��Ȼ���µġ���Щ�����������ӣ����磬���ǿ�Խ�˶��ָй�ģʽ������ͨ����ӳ�˺�������Ҫ���¼���ͳ�����ݣ���ʹ��ѧϰ���̺��ѵ������Ӳ�������ͬ�ı�������ˣ������һ������Ҫ���»���ѧϰϵͳ���ۡ�������������CMSģ�͵���ʽ�ṩ��һ�㣬�ڸ�ģ���У�������Ϊ�ڴ���Ƥ��Ķ���ֲ���֯�ĵط��Է��غͶ����������طţ���ʵ��ѧϰ�ͼ�����¡�
CMS������ļ���ϵͳ��ģ�͵Ľ���ѧ��ͼ����һ������о��ṩ�ģ����о�ʹ�÷�����ά�����������������������������Ƥ���fMRI�����������ݵIJ����ںܴ�̶��Ͽ��������ݶ�ά����������ͼ2a�������ݶ�ά�����Կ���DMN�ṹ��һ���漰�Դ�����������˼ά����һ���漰�����������йٸ�֪�ij����о��������ݶȷ�ӳ����Щ����Ľ����������죨ͼ2b���������������DMN��ñ�����Ϊ�ݶȣ������ǵ������磩��λ�ڸ��ϲ��DMN�����漰����Ϣ�ļ��ɺ�����ʽԽ�ࡣ��С��fMRI�������������п�����ȡ��һ���dz����Ƶ����ݶȡ���Ȥ���ǣ���С���У������������������ݶ��ϵ�λ������Ӻ����嵽���Ա���ľ���������ء���Ҫ���ǣ�DMN�����ж�̬���ԣ����Խ��طŴ���������Ƥ���������λ���ر��ǣ�DMN�����ܹ��ڳ�ʱ��߶��ϲ���������ͼ2c��d�����ٽ�ʱ�����ϣ��Լ��ܹ�ʵ�ֿռ����ϵĴ��ģ�����������ڽ��ٽ�״̬����֪ʶ��1����ͬ�������ǵļ�����ص��ǣ�������Ļ��DMN����Ļ������أ�ͼ2e�����������λ�����ݶȵ�������DMN���Աߣ�ͼ2e����CMSģ�ͣ�ͼ3��������һЩ�ؼ����裬��������һЩ�ɲ��Ե�Ԥ�⣬�������������ۡ�
�ط��������ӽ��䴫����CMSģ�ͱ������ط�����������Ҫ�����ݶȴӺ����崫��������֤��֧���طŴӺ�����DMN�ڵ㴫�����ڲ�ǰ��Ҷ��ѹ��Ƥ�ʺͺ�ҶƤ���������ݶȵ���һ�˷��ֵij����о�Ƥ�ʣ����Ӿ�������Ƥ�ʣ�Ҳ�۲쵽�طš�����Щ�����У��Ѿ���ʾ���뺣���طŻ����ϵ�����Ӻ�������ЩƤ�ʵĶ�ͻ��ͨ·���м䲽���в������Ȼ������ʾƤ�ʻ�뺣��SWR��ص�Ӱ��ѧ�о�ǿ����ʾ�طŻ�����ݶȴ��������⣬MEG�о�����DMN������������Ƥ�����磬�����ܵ����伤�CMSģ��Ԥ�⣬��Ȼ�ط��Ǵ���Ƥ�㣨�����������ձ�������DMN�طŽ��뺣��������أ�DMN����������DMN�����أ��Դ����ơ����仰˵���������ݶ�����Щ�ṹ֮���������ӣ���ͬ�Խṹ���ط�֮�������Խ����ͣ�����������ɼ�¼�������в��ԡ�
������������ڲ�ͬ������CMSģ���У��������DMN�ڵ㣨�Լ��������ݶȵĽڵ㣩�еĻ���ṩ���������룬��Щ�����ʾ�����Լ������εĻ���á�Ϊ��֧����һ���裬���ܴŹ������ʵ�������DMN����Ķ����ػģʽ�����ṩ�й�������Ի״̬����Ϣ��CMSģ�ͱ�������ͬ������������뽫�������Ե����ӣ���Щ���Ա���ͨ����ӹ�������������������Щ������Ҳ������������������ĵͽ�����������������ں���������ָ���Ƕ��ص���ڼ��䣬DMN�����뱻��Ϊ��ӳ�Ը�����֪��DZ��ԭ�������ǵ������У�ͬ��������꣩�ͺ��壨������в����������£�����ʶ��Ҳ����˵�����Ǵ����������ͳ�����ݣ������ӻ���ʾ������Щ��ʾ�ַ�������������������ɲ��洢�ڽϵ�Ƥ�����Ӧ���ԣ�ͬ���Ƥë�������ζ����������������Ļ���Ӧ�ÿ��Ը���һ�������ĻģʽԤ�����������������ݶ��ϵ�״̬�ͻ����Ӧ�ð���Ԥ����������֮��ij�������ԣ����������������ʹ��fMRI�Ͷ����ؽ��뼼�����ڶ�����ʹ�ø߷ֱ������뼼�����в��ԡ�
�ط��ڶ��Ƥ���������������طű���Ϊ���ɻ������ӣ����磬�ں���CA3�����Ŵ�������Է������������������������γɵġ�CMSģ�ͱ������طŵ��������������ں����壬��ʵ֤��������Ƥ��������ȫ���������㹻��������ͨ����֧������ά�ֻ����ˣ����磬����������϶��µ�Ӱ�챻��Ϊ���������ֵ�����ȫ���ط��¼���ԭ������ȷǰ��ҶƤ��Ĵ�����ط��¼�������û����Ӧ�ĺ����ط��¼�������¹۲쵽�ģ����Ӿ�Ƥ����ط��¼�ֻ��һС�����뺣�����¼���һ�¡�û�к������϶���Ӱ����طſ��ܷ�ӳ���ڵͲ㼶������/����ϡ������̵ļ��䣨��ۼ���ǿ����˿��Զ����ں���������¼���������������Ϊ�����������Ӷ����ʱ�����µļ�����Ҫ�����ݶȵĸ���ˮƽ�ϼ����طţ�ͼ3����
����ʹ��ʹ������DMN�ṹʧ��IJ��������Զ������Ȥ�����Է��طŵ�����������CMSģ�ͣ������¼����ط���Ϣ�ĸ����Խ��ܵ����ݶ�������漰�����λ�õ����ơ����⣬���������ݶ��Ͻϵ͵Ľ�С�ط��¼�Ӧ��ӳ�Ͼɵ���Ϣ�����ۺ��Խϲ�����ͨ���Զ����������Ļ���г������������ط���Ϣ��������������������Ϊ�ط��¼����ǵĴ�������Χ�ĺ�����������
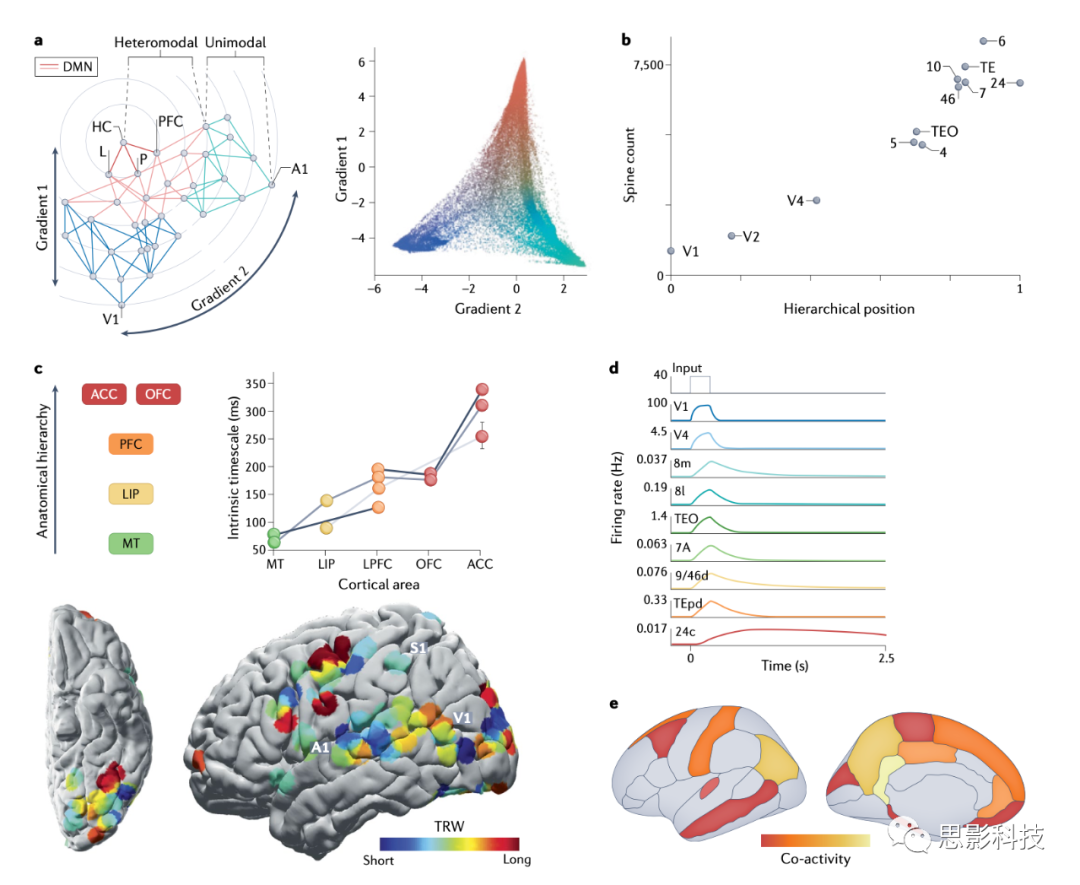
ͼ2 Ƥ���ݶȣ���ͨ�Ժ�����ʱ��߶�
a. һ�ֻ��ڹ�����ϵ��Ƥ���ݶȡ�����ͼ��ʾ���Ժ�Ƥ��ͻ����֯�ķ����ó��������Ľ��ۣ���Ƥ�������Ե��ʾ����ɫ�߱�ʾ����֮��ļٶ����ӣ�������������ֲ�����Ϊ��Ҫ�ݶȣ������ز�ͬ��ͻ��ˮƽ���в���Խ��ͬ��ͻ����ˮƽ����ͬ�Ļ���ʾ��������Ϊ��Ϣ�Ӹо��������㵽������������ͻ������Ծ�����������ֶ�ά���н���������dz��ɫ������ɫ�����еļ�϶��ʾ��ͬ�й�ģʽ֮��ı߽磩�������ݶ�1��һ�ˣ����������ɫ����Ĭ��ģʽ���磨DMN�����ص��������ڸ��ݶȵ���һ�ˣ��Ӷ��γ��˼ӹ��εIJ�νṹ�����ǽ��飬�����������Ϊλ��DMN�Ϸ����ݶ�1��εĶ��㡣�Ҳ���ʾ�˾�Ϣ��������Ԫ������ʾ�ĵ�Ч��֯ԭ�����ݷ�����Ƕ��ά��������õ�ǰ����ά�Ȼ���ʱ��Ƥ�����أ��ɲ�ɫ���ʾ�������������ʾ��ͬ���ݶȾۼ��������ݶ�1�ڳ����о��˶�������ɫ���Ϳ��������ɫ��֮�����죬�������ݶ�2�������˶�������Ƥ�ʣ�dz��ɫ�����Ӿ�Ƥ�ʣ�����ɫ���ֿ���
b . �ں����У�Ƥ������ϸ���ϵ���ͻ���������Ÿ�������Ƥ�ʲ�νṹ�е�λ�ö����ӣ�����ͨ�������ٷ����ó��ģ�������a������ʾ�����ֱ�ǩָ⨺�����еIJ�����������
c .����ѧ��ʱ��߶�����Ƥ���νṹ�������Ͻ������ʾ���Ӹо��������DMN��������������ӡ����Ͻ������ʾ�����ӵĵ�����Ԫ�������ʱ��߶������Ӿ�-ǰ��Ҷ��νṹ�����ӣ�ÿ����һ��������ʵ�����ݼ���������������ʾ������������������ʹ��Ƥ�����������������Ҷ�߶ȡ�ʱ����ܴ���TRW������Ԫ����������Ϣ�Բ����ɿ���Ӧ��ʱ�䳤�ȵ�һ�ֶ��������ڳ����о�����������������Ƥ�㣨A1���������Ӿ�Ƥ�㣨V1���ͳ�������о�Ƥ�㣨S1����������̣����䳤��Զ��о���������Ƥ������������������ģ�Ͳ�����һ�����Ƶ���ʱ��߶������ס���ʾ��ģ����Ƥ�������V1�������������Ӧ���ڸ�ģ���У����������Ǹ��ݺ������о��л�õ����ӷ����������еģ��������������ģ�ͻ��Ľ��ʲ�δ���ʱ��Ƥ���������Ӧ��ʾ��Խ��Խ����˥��ʱ�䡣����ʾ�˴ӵײ�V1��������ǰ�����Ĺ��ܲ�νṹ��������DMN�����γ�ǿ��Ĺ�����ϵ����ʾ��ͼ�ϣ���ɫ��ʾ�ھ�Ϣ״̬�£��������뺣������ֳ��ɿ�Эͬ����������۴��ء�ШǰҶ���ڲ��Ҷ����������ʾ�����
ACC��ǰ�۴���Ƥ�ʣ�HC��������L,��ԵƤ�ʣ�LIP���ඥҶƤ�ʣ�LPFC����ǰ��ҶƤ�ʣ�MT���������OFC �ۿ�ǰ��Ƥ�ʣ�P����ҶƤ�ʣ�PFC��ǰ��ҶƤ�ʣ�TE�����ǰƤ�ʣ�TEO����º�Ƥ�ʣ�TEpd�������Ƥ�ʡ�
�ֲ������϶��º����¶��϶��Է����Ӱ�졣�ڴ��Ե�ÿ����λ��CMSģ�ͱ����Է����ӳ��������ͬ�����أ��ֲ������Ļ����������������������¼�������Ը����������Ӱ�죨���������������������ԣ����������ݶȸ����µ������Ӱ�������Դ�����ģʽ��ɣ��Ӷ������ֲ����������ϸ�����£����ֲ����ɵļ�����������ϱ��ؼ��䣬�������κ����η�����Ϣ�����϶��µ�Ӱ����������������ӣ������������������ָ�Ľϵͼ������֮�佨�����ӡ����¶��ϵ�Ӱ����ܻ�ʹ������������¼������ƫ��Ӷ��ṩ��ѧϰ�������õķ�������Щ������ܻᱻ���ԣ����磬�����λ���������ʧ��ʱ�������ڸ��������й۲쵽�طţ����ҿ��Ա���ͳ����������Ϣ���ݵ���Ӧ�仯��������Ϊ��ͣ����Ŧ�����ƻ����ε���֮�����ͨ�ԡ����磬������ʧ����ܻ���ֹ����Ƥ���п�ʼ���ط��¼��������Ӿ�Ƥ�㣬����ֹ�¼�����Ķ�ģ̬���ϡ���������ļ�����˵��Ч��Ӧ�ø�ǿ��DMN�����ʧ���������Ƶ�Ч���������ϵļ����и�ǿ��Ӱ�졣
ȫ���طŲ�ȡ��ʱ�㷺�������ʽ��CMSģ��������ط���ؼ�����ѩ����ʽ����������ʾ���ٽ������ͳ�����ݣ�ѩ����С�����ɷֲ���֪ʶ��1����������ǿ����Ϣ�Ӻ�������Ƥ�ʵ��������������෴��CMSģ����Ϊ�������ṹ�γ���һ���Է���Ϊ����ϵͳ�������ں�����䷴��һ������㷺�Ļ������ijЩ����£��漰�����ԵĴ֡�����ͨ���ڽ��ٽ�״̬�¹���������������ʶȵ���������ӣ�֪ʶ��1�����Ӷ�����ȵ����Զ������ԣ����Ŵ�ңԶ����֮��ϡ��Ķ�ͻ�����ӡ�ͨ�����ְ��ţ�ѩ�����ܻ������ݶ�������Զ������֮���������ԣ��Ӷ�������������֮������ӣ�����Щ������������νṹ�в���ֱ�ӵġ��ϼ������¼����������Ʋ⣬����ܵ��¸��ӱ���ѧϰ�Ĵ��ģ���١����ݶȽϸߵ����������������ϴ��ѩ�����翼�ǽ�����ͨ�Ժͼ��ܶȵļ����о���ʾ��ͼ2b����Ϊ�˲���ѩ����ѧϰ�ͼ����е����ã��������롰�ٽ�㡱�������Զ�Ĵ���������ݻ�����磬ͨ���ջ��̼����������Լ��乮�̺����Ӱ�졣
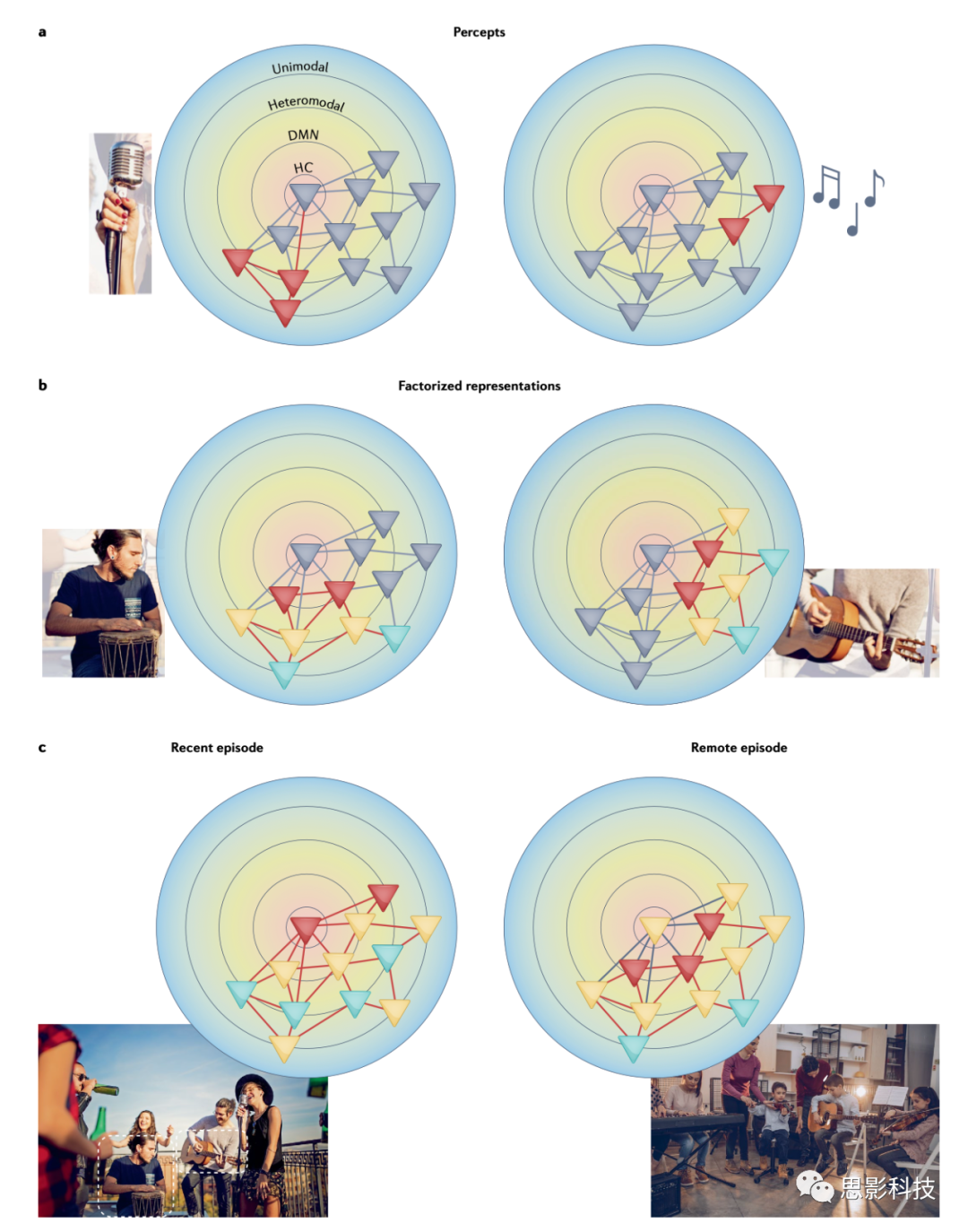
ͼ3 CMSģ���е�Ƥ�㼤��ģʽ��
CMSģ�͵�ʾ��ͼ������Ƥ�������Էֲ���ʽ��ʾ����ǰ��������ͼ2������ɫ��������˴�Ĭ��ģʽ���磨DMN������ɫ��������о�������ɫ������Ҫ���������ݶȣ��Լ�DMN�Ϸ��ĺ����ṹ����ɫ���������α�ʾ������������ɫ��ʾ��˲̬�¼��ڼ䱻���輤���˳��-��-�̣�����ɫ�����α�ʾδ������������ߺͻ��߷ֱ��ʾ˲̬�¼��ڼ伤���δ���������·����
a. ���ڼ�֪��������۲쵽���ݵľֲ�����漰�������������������磬�ڸ��ֵ��֣������ֵ�������֪���ң�������£����ܻᷢ�������������Щ���ݵļ��������ǿ��ľֲ��ظ�����֧�֡�
b . ��ʱ���������DMN���������������ݶȵ�DMN�������������빹���¼��Ķ���ԭ������صı����ģ��٣�����������ڼ�����ʱ�����Ӧ����Щ��������������б����������������������ǰ��õ�֪ʶ�����磬������������ߣ���ͼ���ı��ֿ��ܰ����������ݡ�����Ľ����Լ������ֶ�������Ա�Ĺ�ϵ����Ϣ����һ��Ƥ�����磨��DMN�������һ���Ӽ�����������������ļ��������������״����������������������ԡ�
c . ����CMSģ�ͣ������������������ڣ�������һ�������ɶԣ��������������Ϻͼ�����ȫ��Ƥ��˲ʱ����֧�֣�������������£��������������HC����������������˲���������ؼ����á�����Զ����ڣ������Ƕ�ͯ���ֿΣ��������ܿ�����DMN�ṹ�����������ܻἤ��������뻥��ѧϰϵͳ�������ƣ����ּ��輤���Ƕ��ݵģ�ȡ���ڴ���Ƥ��Ľ��ٽ綯�����ƣ����ɺ�����Ϊ���ڼ���������DMN����ΪԶ�����������������ˣ������ģ���У���ں�������������ͬ�Ķ�̬����֧�֣�����Ӧ�ڲ�ͬ�߶ȵļ��
����
����CMS������ļ���ϵͳ�������ۺ��˴����۲������γ����µĹ��ܹ۵㡣���ṩ��һ��������������ȫ����Ի�Ĵ��ģ�������Լ���ϸ���ֱ������ݣ������磬�����Խ���ȫ�Կռ�������ɺ���Э��������β����ģ��Լ������о�Ƥ���г��ֵĻ��ѡ���Ա��������á����⣬�������Խ�����������ij��֣��������������ģ�⸴������Ļ�����������п��ܴٽ�����������ݣ�ϸ�������븴����֪��ս���ϵ����ݺ�����ã��붯�����ݣ��۲����������Ϣ�������ռ���֮���Э����
�����ԣ�CMSģ�������������裬��Щ���赼��Ԥ����Ȼ��Ҫ����ʵ����Ժ��������ۼ��顣����ʵ�齫�ǹؼ������ȣ�����ʧ��ͶϿ�ʵ����������о��طŹ��������¶��Ϻ����϶��µ���Ϣ�����������Ҫ���ǽ���BOLD�ź�ˮƽ�ϻ�õ����ݣ�����DMN�����ʹ�õ�����ԪȺ��¼��������ϵ��������ȷ��DMN�еĹ�ͬ�����Ƿ�ӳ���ٲ����ɺ���Э�����طţ��Լ�����Ϣ�Ƿ�������Ƥ�������硣���������Ȼ�����ѣ������������ṩ��ȷ�Ľ����Ȼ�����漰���ģʽ���ۺͽ�ۣ���ʵ�飨���磬ʹ�ý��뼼�������ӡ����ࡱ����Ļ���ܻ���������⡣�������������ۼҺ�ʵ����������˵��һ�������ǣ�����ѧ�����Ĺؼ������Ƿ����������ѧϰ���̡���һЩģ���Ѿ���ʼ�����һ�㣬�����ڴ����ٽ�״̬��ʵ�飬�����Ǿֲ�����ȫ�֣�������������֧�ֻ��Ʒ���һ�۵㡣
CMSģ��Ҳ��һЩ���ޡ����磬���Ѵ��Ե�Ƥ�㡰ȥͬ����״̬��˯�ߴ��Ե��ٽ�״̬��ͬ����ˣ����ǵ������Ƿ�Ҳ����������״̬�������䱻����ʱ���в�������ٽ�״̬������Ȼ����������״̬���������ڶ����¼�����Ԫ�Ӽ������߲�ͬ�Ķ���ѧԭ�����ڷ������á����⣬�������⣬DMN�ڼƻ���ģ�ⷽ��Ҳ����Ҫ���ã���Щ���̿����漰�����طţ�����2������Ȼ�����طŶ���Щ���ܵĹ������ۿ��������ǵļ���ģ��������أ���Ҳ������Ҫ�����ش������
�ܽ���ԣ����ǽ������о�������ϵ�����������������ںܴ�̶����Ƿֿ����еģ������˲�ͬ�����ֺͼ�������ij�̶ֳ�����ѭ�˲�ͬ�����۷���ͨ��ǿ��DMN���طŹ����д��ڵĽ���ѧ�����ܺͻ����ѧ�Ĺ��ԣ�����ϣ�������µļ��⣬������Щ����֮������εĻ������Դﵽ���䡢�ƻ���ģ����̵�ͳһ���ơ�
����ԭ�ļ��������������˼Ӱ�Ƽ��ţ�siyingyxf��18983979082��ȡ,���˼Ӱ�γ̼��������ȤҲ�ɼӴ��ź���ѯ����˼Ӱ�ṩ����������ط�������ҪҲ�����Ӵ��ź���Ⱥ��ԭ��Ҳ����Ⱥ�����������ǵĽ���������о��а����������ת��֧���Լ����½ǵ��һ���ڿ����Ƕ�˼Ӱ�Ƽ���֧�֣���л��

��ɨ����߳���ѡ��ʶ���ע˼Ӱ
�dz���лת��֧�����Ƽ�
��ӭ���˼Ӱ�����ݴ���ҵ�γ̽��ܡ�����ֱ�ӵ���������ּ������˼Ӱ�Ƽ����еĿγ̣���ӭ�����ź�siyingyxf��18983979082������ѯ�����пγ̾����ű��������������ǻ��һʱ����ϵ���������ѱ���ѧԱ�����
�˴ţ�
�Ͼ���
����ʮ����Ź������������ݴ����ࣨ�Ͼ���10.16-21��
�ڶ�ʮ����Ź�����Ӱ��ṹ�ࣨ�Ͼ���10.24-29��
����ʮһ��Ź�����Ӱ������ࣨ�Ͼ���11.12-17��
�Ϻ���
�ڶ�ʮ�Ľ���Ӱ�����ѧϰ�ࣨ�Ϻ���10.9-14��
����ʮһ��Ź������������ݴ����ࣨ�Ϻ���10.28-11.2��
����ʮ�Ž�Ź�����Ӱ������ࣨ�Ϻ���11.4-9��
���죺
�ھŽ����������ݴ�����߰ࣨ���죬10.13-18��
����ʮ��Ź�����Ӱ������ࣨ���죬10.22-27��
�ڶ�ʮ�˽���ɢ�������ݴ����ࣨ���죬11.5-10��
��������ɢ�Ź��������߰ࣨ���죬11.17-22��
�ڶ�ʮ����Ź�����Ӱ��ṹ�ࣨ���죬11.27-12.2��
������
��ʮ�����������ݴ�����߰ࣨ������10.20-25��
��ʮһ��Ź���ASL������������ǣ����ݴ����ࣨ������11.3-6��
����ʮ����Ź�����Ӱ������ࣨ������11.9-14��
������Ӱ����ѧ�ࣨ������11.25-30��
�����R����ͳ�ưࣨ������11.16-20��
�Ե缰���⡢�۶���
�Ϻ���
�ڶ�ʮ���������Թ������ݴ����ࣨ�Ϻ���10.17-22��
����ʮ�����Ե����ݴ����м��ࣨ�Ϻ���11.13-18��
�ڶ�ʮ�˽��Ե����ݴ������Űࣨ�Ϻ���11.20-25��
������
����ʮ�Ž��Ե����ݴ����м��ࣨ������10.11-16��
���£���ʮ�����۶����ݴ����ࣨ������10.26-31��
�Ͼ���
������Ե����ѧϰ���ݴ����ࣨMatlab�汾���Ͼ���11.3-8��
���죺
�ڶ�ʮ�߽��Ե����ݴ������Űࣨ���죬10.28-11.2��
���ݴ���ҵ����ܣ�
˼Ӱ�Ƽ����ܴŹ���(fMRI)���ݴ���ҵ��
˼Ӱ�Ƽ���ɢ��Ȩ����DWI/dMRI�����ݴ���
˼Ӱ�Ƽ��Խṹ�Ź����������ݴ���ҵ����T1)
˼Ӱ�������������У�QSM)���ݴ���ҵ��
˼Ӱ�Ƽ������ද���С����Ӱ�����ݴ���ҵ��
˼Ӱ�Ƽ��鳤�ද��fMRI����ҵ��
˼Ӱ���ݴ���ҵ������ASL���ݴ���
˼Ӱ�Ƽ���Ӱ�����ѧϰ���ݴ���ҵ�����
˼Ӱ�Ƽ������Ⱥ����ҵ��
˼Ӱ�Ƽ�EEG/ERP���ݴ���ҵ��
˼Ӱ�Ƽ��������Թ������ݴ�������
˼Ӱ�Ƽ��Ե����ѧϰ���ݴ���ҵ��
˼Ӱ���ݴ������������Դ�ͼ��MEG�����ݴ���
˼Ӱ�Ƽ��۶����ݴ�������
��Ƹ����Ʒ��
˼Ӱ�Ƽ���Ƹ���ݴ�������ʦ ���Ϻ����������Ͼ������죩
BIOSEMI�Ե�ϵͳ����
Ŀ��ʽ���ܴŹ���̼�ϵͳ����