由德国亚琛工业大学医学中心团队主导的一项近红外超扫描研究即将发表在NeuroImage期刊上。亲子同步在社交接触过程中行为和生物信号的耦合可能会影响儿童的情绪纽带形成以及和儿童情绪调节发展相关的大脑回路。本研究通过测量父母和孩子的前额神经活动探究这一过程的神经生物学基础。每个孩子分别与父母(主要是母亲)以及成年陌生人一起玩合作游戏和竞争游戏。结果发现在合作期间,父母和孩子的大脑活动在DLPFC和额极皮层(FPC)出现同步,其可以预测实验中的合作表现。在亲子竞争(CoopP)、陌生人与儿童合作(CoopStr)以及陌生人与儿童竞争(CompStr)的三种条件下,均没有观察到明显的脑同步。此外,在合作情境中,和陌生成人小孩组相比,亲子之间的脑同步对父母情绪调节能力和儿童情绪调节能力之间的关系有中介作用,这一结果和问卷评估结果一致。因此,该研究认为:脑同步可能代表着父母与孩子之间情感联系的潜在神经机制,它与儿童适应性情绪调节的发展有关。未来的研究可以从以下角度开展:脑同步是否可以作为双人社会-情感交互的神经生物学标志,并且是否可以通过干预来改善。
关键词:近红外 超扫描 亲子互动同步性
研究方法
被试
共有33组年龄在5岁至9岁(M = 7.52岁,SD = 0.87岁)的儿童与父母一起参加了这项研究。其中包括13个母女两人组,17个母子两人组,1个父亲女儿二人组,以及2个父子二人组。此外,成年陌生人与参与的孩子(小孩 - 陌生人两人组)完成相同的任务。研究中共有21位成年人担任陌生人:其中,15位陌生人参加过一次,2位陌生人参加了两次,3位陌生人参加了三次,1位陌生人参加了四次(更多信息参见文本A.1 )。陌生人性别和父母性别进行匹配;父母的年龄显著大于陌生人(父母:M = 41.24岁,SD = 4.32岁,范围:33-49岁;陌生人:M = 24.33岁,SD = 4.70岁,范围:20-45岁; t (32)= 15.74,p <0.001)。最终,7个fNIRS数据由于孩子不顺从实验指导或技术原因而丢失(4x CompStr,2x CoopStr,1x CompP)。
亲子被试通过以前RWTH亚琛大学医院的研究招募。陌生人通过研究人员和被试的社交网络招募。支付参加研究的家长、儿童和成人陌生人的费用。
实验工具
电脑游戏任务。实验中使用的计算机游戏任务由崔等人开发(2012年)并修改为适合儿童参与的;每个玩家使用按键操纵海豚在屏幕上的移动,其目的是一起抓到球(合作任务)或为自己赢得球(竞争任务)。
在合作任务中,要求被试尽可能同时按键以“一起接球”。如果两人的反应时之差低于阈值,则两只海豚跳到球上(反馈屏幕),抓住球(结果屏幕)并获得一分。如果两人的反应时之差超过阈值,只有更快的海豚才能跳到球上(反馈屏幕),海豚既不能抓住球(结果屏幕),还会失去一分。被试可以根据反馈调整自己的反应时。阈值设置为T = 1/8(RT1 + RT2),其中RT1和RT2是两名被试的反应时。
在竞争任务中,要求被试比其搭档更快做出反应,以“抓住并赢得球”。只有更快的海豚跳到球上(反馈屏幕)并抓住球(结果屏幕)。在每一个试次中,更快的海豚可以获得1分,而较慢的海豚则失去1分。如果两名被试同时以50毫秒的误差幅度作出反应,那么两个海豚都赢得了一分。
实验程序。
被试在进行合作或竞争游戏时并排坐在一起。要求其在fNIRS测量期间保持头不动,并避免与其他玩家对话。将毛巾放在双方手上以减少被试观察彼此活动的能力。在合作和竞争任务开始时先进行五次练习试验。任务流程如图1所示,在每一个trial中,屏幕上均显示两只海豚。2秒后,两只海豚上方出现一个空心的黑色圆圈(“准备好”信号),并在0.6-1.5秒的随机延迟后保持在屏幕上,直到它被一个彩色球(“去”信号)代替。要求被试只在“Go”信号出现后再去按键。在被试较快反应(合作/竞争)之后,呈现反馈屏幕,再显示结果屏幕。反馈和结果屏幕均在竞赛条件任务中呈现1 s,合作任务条件中呈现1.5 s。
设计。
每个计算机游戏任务由两个任务块组成,每个block包含20个试次和3个30秒静息时间,按以下顺序:休息,任务块1,休息,任务块2,休息。在儿童之间平衡两种电脑游戏任务(合作/竞赛)的顺序,但是保持两人组不变。14个孩子首先与父母一起完成这两项任务,在短暂休息后接着与陌生人完成这两项任务。而另外19个孩子则颠倒顺序进行电脑游戏任务。
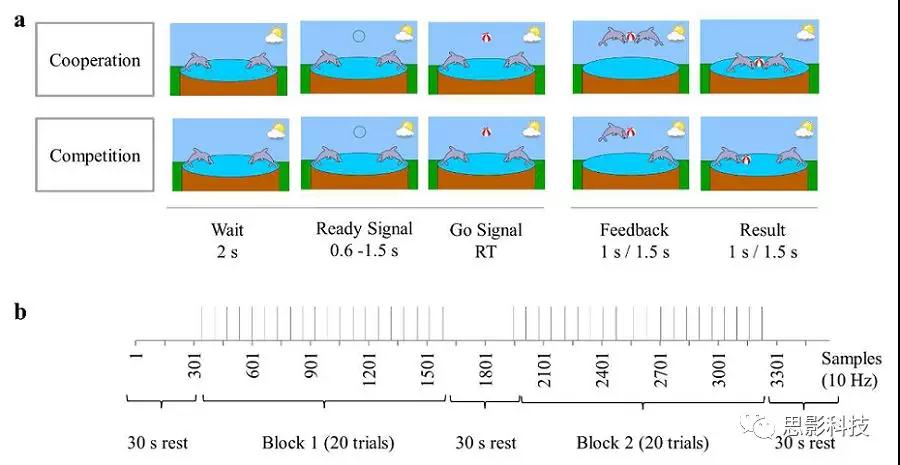
图1.实验设计图(a)合作和竞赛电脑游戏任务期间的个人界面。任务是尽可能同时对“Go”信号(合作)做出反应,或者比合作伙伴(竞争对手)做出更快的反应。反馈和结果屏幕在竞争游戏中出现1秒,合作游戏中每次出现1.5秒。 RT:较慢被试(合作)/较快被试(竞争)的反应时间。(b)电脑游戏任务全观。单个试次可视为垂直线。合作和竞争均由两个随后的任务块组成,每个块有20个试次,相隔30 s的休息时间。
对任务表现的行为测量
在合作和竞争任务期间记录两名被试的反应时以及反应时之差。由于数据中存在异常值(>3 SD),因此分别报告每名被试和二人中位反应时(中位数RT)和反应时间差(中位数-DRT)的中位数(根据Panet ,2017年)。为了进一步量化被试的任务表现,报告合作期间共同获胜的比例以及竞争期间儿童获胜的比例。
情绪调节测量
情绪调节清单。情绪调节清单(ERC)(Shields and Cicchetti 1997)是一个具有24个题目的问卷,使用4点李克特量表(1 =几乎一直到4 =从不)来测量与孩子的情感不稳定性,强度,效价,灵活性和情境适当性有关的过程。问卷由父母填写。根据Shields和Cicchetti(1997)的描述计算复合情绪调节分数,分数越高表明孩子的情绪调节能力越好(当前样本中的克伦巴赫α= 0.74)。样本项目包括“可以说当他/她感到伤心,生气或愤怒,害怕或恐惧”以及“可以从不安或痛苦的情境中快速恢复”。
fNIRS数据获得方法
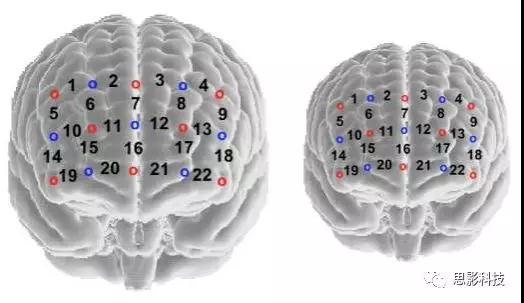
使用单个NIRS设备(ETG-4000,日本日立医疗公司)来同时测量两名被试氧合血红蛋白(oxy-Hb)和脱氧血红蛋白(deoxy-Hb)的浓度变化。采样频率为10 Hz。将一个“3×5”光极板连接到由坚固织物制成的盖帽上(Easycap GmbH Germany)。在每个贴片中,定位了八个发射器和七个接收器,从而产生22个测量通道(CH)。源探测器距离为3厘米。如Cui等人所述,将帽子对称放置在参与者的额头上(2012年),以便帽子的底部触及被试眉毛的顶部,中间探针列在鼻子上方对齐。
空间定位(重要!)
为了估计通道的空间位置,研究者将独立fNIRS数据的虚拟注册结果用于立体定向脑坐标系统(Singh et al.2005; Tsuzuki et al.2007)和自动化的Talairach地图集标签(Lancaster et al。2000)。光电极的位置与日本自治大学提供虚拟注册结果的“3×5”固定器位置之一非常相似:最低探头行的中间光极放置在极点中点(Fpz)上,中间探针列沿着矢状参考曲线对齐(http://www.jichi.ac.jp/brainlab/virtual_registration/Result3x5_E.html)(图2)。
fNIRS数据预处理
使用SPM_fNIRS工具包进行信号预处理(https://www.nitrc.org/projects/spm_fnirs/)。将修改后的Beer-Lambert定律用于计算氧合血红蛋白和脱氧血红蛋白的变化。差分路径长度因子根据波长和被试的个体年龄来计算(Scholkmann and Wolf 2013)。运用略微修改以后的自带MARA算法(Scholkmann et al。2010)来减少运动伪迹。此外,使用基于离散余弦变换集的高通滤波器(SPM12 6906版本,http://www.fil.ion.ucl.ac.uk/spm/software/spm12/)减少低频噪声。噪音过高的通道通过肉眼检查识别,将其排除在进一步的分析之外(大约9%的通道)。
脑同步分析
小波相干分析。根据以前的研究(Cui et al., 2012; Cheng et al., 2015; Baker et al., 2016),计算了相应通道的oxy-Hb时间序列之间的小波相干性作为衡量脑间同步性的指标。所有的小波分析均采用ASToolbox(Aguiar , Conraria and Soares 2014)进行,如下所述。我们选择了广义Morse小波(GMW)(Lilly和Olhede 2012),其参数β= 3和γ= 3。通过改变这些参数,GMW可以采取多种形式。对于γ= 3,与其他整数值γ相比,小波是最对称的,最接近高斯的,并且大多数时间频率集中,而保持完全分析(小波也被称为“艾里小波”)。
显著相干性。为了估计二维矩阵中每个小波相干值的统计显著性,使用AR(1)模型拟oxy-Hb信号,并且使用自举法构建300个替代信号(Aguiar,Conraria和Soares 2014)。如果在特定的时间和规模观察到的相干性值高于在同一时间和规模上获得的替代信号的相干性值的99%,则认为它是显著的。根据以上分析,得到0.65的截断值,相干性值超过该值便达到统计显著性。
任务相关频段。研究者首先确定了一个和任务相关的相干性可能发生的频段,该频段在0.5 Hz和0.08 Hz之间(周期长度为2.02 s - 12.80 s)。根据试验持续时间,以前的研究和对数据的视觉检查来选择该频段。
结果测量。虽然以前的研究计算了小波相干性的平均值,但本研究报告小波相干性数值高于在与任务相关的频带中的截断值的值,将其除以该频带中小波相干值的总数。该比率分别使用block1和block2中的小波相干性计算得到。随后,将这两个值分别进行平均,以产生一个用于合作条件下的一致性值(Coherence),以及一个用于竞争条件下的一致性值。这个比例被用作脑间同步的主要结果变量,并且在下文中用“一致性”表示。另外,本研究直接比较了四个条件(CoopP,CompP, CoopStr和CompStr)的一致性。研究者通过减去各自的一致性,计算了亲子合作与亲子竞争(CoopP-CompP)的一致性以及亲子合作与陌生儿童合作(CoopP-CoopStr)的一致性的个体对比分数值。这样进行比较的原因是过去的神经影像学研究结果表明适当的任务条件通常比休息条件提供更好的基线(Stark and Squire 2001)。
随机配对分析
单被试的FNIRS信号可能由于相同的实验条件(检查假双人组的EEG同步:Burgess 2013)和常见的缓慢生理振荡(如呼吸和脉动)而显示相似性。因此研究者进行了旨在揭示与实验交互无关的脑同步验证方法。在验证程序中使用随机对(随机对指的是双方不是同时进行实验而是执行相同的实验任务的亲子对的小波相干性)分析方法,计算随机对并将其与相应通道中的实际交互对的小波相干性进行比较。应用“数据分析”中描述的一系列t检验来表明实际配对中的相干性是否显著高于每个通道和条件的随机配对。虽然这种通过随机配对分析的验证程序已经可以提供关于实验条件之间潜在差异的初步方法,但它不能代替直接统计比较。此后的分析只考虑至少一种实验条件下相干性显著高于随机对的相干性的通道。通过排除那些没有显示有效相干性的通道(与随机水平以上的交互有关),研究者降低了虚假风险结果,从而提高结果的可靠性。验证分析中对1200个随机对(四个条件中的每个条件为300个随机对)计算小波相干性。噪音过高的通道被排除在分析之外,因此随机对相干值的数量在通道之间变化。应该注意的是,即使两个被试执行相同的实验任务,由于反应时间和可变的实验间隔的差异,block的时间和长度也不相同。由于相干性分析要求两个时间序列的持续时间相等,因此在最后分割较长的block以便它与每个随机对的较短block的长度相同。对于所有block分别重复该过程。之后,将它们重新连接起来,计算整个Oxy-Hb时间序列的小波相干性。
数据分析
使用IBM SPSS Statistics 22和R 3.4.1(R Core Team,2017)进行统计分析。所有的统计检验都是双尾,95%置信区间,通过bootstrapping(n = 5000次检测)得出。为了检验双人对的相干性值是否显著高于随机成人孩子对,进行了独立样本t检验和Welch's t检验。如上所述(“通过随机对分析进行验证”),所有后续分析仅针对显示出比随机对的相干性显著更高的通道进行。
为了直接检验不同条件之间脑与脑相干性的差异,研究者将各自通道中具有相干性的线性混合模型(LMM)用作因变量。为了探究影响因素,儿童的年龄(以月为单位),儿童的性别以及条件顺序分别添加到LMM。LMMs用R包lme4进行(Bates等,2015)。对于统计推断,模型使用REML拟合,p值由使用Satterthwaite逼近自由度的lmerTest软件包的功能summary / anova推导。
使用配对样本t检验探究行为测量中的条件差异。行为测量与相干性之间的关系通过控制孩子性别的偏相关、Pearson相关性以及等级多重回归来检验。
情绪调节以及相干性之间的关系通过控制孩子性别的皮尔逊相关来检验。之后,用SPSS的PROCESS宏进行调解分析(Hayes 2013)。通过95%置信区间的偏差校正自举法(n = 5000次运行)测试了间接效应。
由于多次检验,研究者使用FDR程序校正p值。对于随机对的比较,校正的t检验数为88(对应于四种条件下的22个CH)。 LMMs的p值在两次检验中均被调整,因为LMMs是针对两个通道进行的。通过检验四种简单效应来分解双向交互作用,因此p值被调整为四个检验。对于任务表现的相干性和行为测量之间的关系,校正的检验数量是4个(四个相关和四个回归)而对于相关性和情绪调节之间的相关性数量是18个。
结果部分
亲子合作条件下的同步性
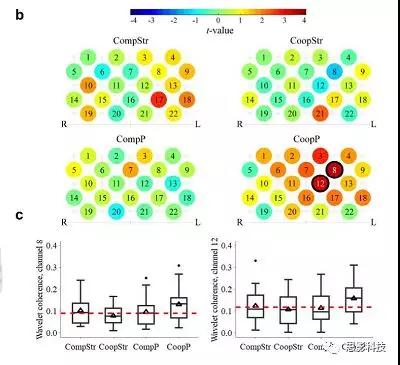
随机对之间比较的结果。为了识别相干性增强的通道,研究者比较了双人组的相干性和随机成人孩子对的相干性,结果如图2所示,在合作过程中亲子的相干性(CoopP)在CH8(t(946)=3.48,p<0.001,padj.=0.038,dCohen= 0.67)和CH12中(t(1017)=3.34,p<0.001,padj。=0.038,dCohen= 0.63)显著高于随机陌生人儿童对,这些通道对应于背外侧前额叶皮层(DLPFC;布罗德曼区,BA9)和额极皮层(FPC;BA 10)。在经过FDR校正以后,没有任何通道在任何其他情况下(CompP,CompStr或CoopStr)显示出显著的相干性。
实验条件之间的比较。
为了直接比较实验条件之间的差异,研究者在CH8和CH12中使用LMMs考察了任务(0=竞争;1 =合作)和实验搭档(0=陌生人,1=父母)相干性的主效应和交互效应。对于这两个CH,任务或搭档主效应不显著;但是二者的交互作用显著(LMM的结果显示在表1以及图2的箱线图中)。即如固定效应估计所示,CoopP条件下的相干性增加。该结果得到了简单效应分析的进一步证实,其结果发现只有在亲子合作任务条件下才具有更高的相干性(CH8:t(27.48)=2.11,p= 0.044,padj.=0.088; CH 12:t27.29)=3.06,p= 0.005,padj.=0.020)。而经过FDR校正以后只有在CH12中出现这种显著影响,而CH8边缘显著。相应地,实验搭档仅对合作任务有显著影响,两个CH中的亲子的相干性更高(CH8:t(51)=3.21,p= 0.002,padj.=0.009; CH 12:t(56)=2.53,p= 0.014,padj。= 0.029)。
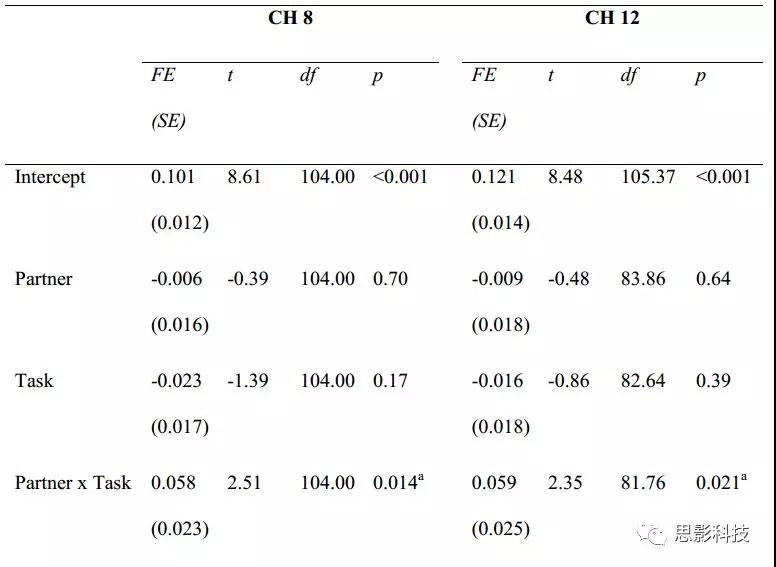
表1. N = 33(CH 8中108个观测值,CH 12中观测113个观测值)。FE(SE)=固定
效果估计(标准误差)。 t = t检验使用Satterthwaite近似来表示自由度。 intercept是指在陌生儿童竞争条件下的相干性。根据FDR校正后的α水平呈显著性(阈值:p≤0.021)
影响因素。为了确定使得CoopP条件下相干性增加的影响因素,研究者分析了儿童的年龄和性别以及实验条件顺序对CH8和CH12相干性的影响。儿童的年龄和实验条件顺序对任务x伙伴交互作用没有显著影响(三因素交互作用不显著,p>0.48)。然而,结果显示CoopP条件下相干性的增加主要是由女性儿童造成的,这些女孩主要与母亲一起参与:任务,实验搭档和儿童性别之间的交互作用显著(CH8:t(100)=- 2.04,p= 0.044; CH12:t(78.8)=-2.13,p= 0.037)。
总之,这些结果表明,只有在FPC和DLPFC中,亲子合作(任务x实验搭档交互)期间,脑间同步才会增加。随后的分析需要考虑到性别差异,而儿童的年龄和实验条件顺序并没有系统地影响任务和实验搭档的交互。
亲子合作条件下同步性的提高不能归因于任务行为的差异
为了检验CoopP条件下增加的相干性是否归因于被试的行为差异,研究者比较了不同条件之间被试的行为差异,行为相似性和任务表现。行为相似性通过双人组在反应时之差上的中位数(中值-DRT)来测量。该指数在竞争条件下显著小于合作条件,这表明竞争期间亲子(t(30)=6.21,p<0.001,dCohen= 1.25)和陌生儿童对(t(27)=4.24 ,p<0.001,dCohen= 1.07)的行为相似性。亲子对和陌生儿童对之间在合作条件下(t(29)=1.47,p= 0.15)或竞争条件下(t(26)=0.70,p= 0.49)没有观察到中值DRT的显著组间差异。
使用合作期间双方均得分的相对试次数目以及竞争条件下儿童得分的相对试次数目来衡量任务表现。亲子的合作率没有显著差异(t(29)=-0.69,p= 0.49)。然而,在竞争条件期间,与陌生人条件相比,孩子更经常地赢得父母(t(26)=5.55,p<0.001,dCohen= 1.08)。综合起来看,这些结果表明,合作期间观察到亲子脑间同步性的增加并不是由可观察到的行为上的差异导致。
亲子合作条件下同步性的提高预示着更好的合作表现
接下来,研究者分析了通道8和通道12在CoopP条件下的相干性是否与合作表现有关。虽然在前面的分析中,研究者将双人在两个合作block中的平均一致性作为结果衡量,现在分别研究每一个block的相关性。首先,分析了CoopP条件下的相干性与每个block内的合作率之间的关系。结果表明,无论在方框1(CH8:pr= 0.32,p= 0.11; CH12:pr= 0.10,p= 0.61)还是在方框2中,相干性与控制孩子性别以后的合作率都没有显著相关(CH8:pr= -0.13,p= 0.54; CH 12:pr= -0.10,p= 0.60)。
其次,应用分层多元回归来检验CoopP条件预测了两个block之间的合作表现,反之亦然。结果表明在block1中,控制儿童的性别和双人在block1中的合作率,结果发现CH8和CH12的CoopP相干性可以显著预测block2中的CoopP合作率(CH8:F(3,23)=5.66,p= 0.005,padj.=0.019; CH 12:F(3,25)=4.56,p= 0.011,padj.= 0.022;表2)。相反,CoopP合作率在block1中不能显著预测block2中的CoopP相干性。因此,在FPC和DLPFC中增加的亲子脑同步性似乎可以使之产生更好的合作表现,而合作表现并不影响脑同步。
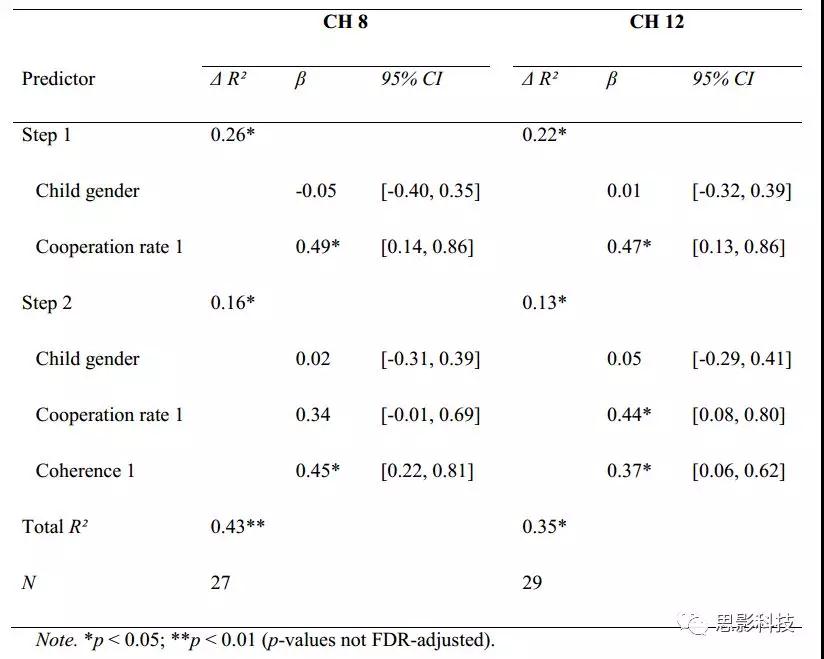
表2.在方框2中预测亲子合作率的等级多元回归分析结果
亲子合作条件下的同步性增强与父母和子女情绪调节有关
父母和孩子的情绪调节。ERC综合评分测量的儿童情绪调节与ERQ测量呈显著相关(pr=0.42,p= 0.017,95%CI= [0.13,0.68];图A.2a)。越多父母使用重新评估作为调节自己情绪的策略,孩子的情绪调节能力越好。父母抑制和儿童情绪调节没有显著相关(pr= 0.04,p= 0.82)。此外,儿童的情绪调节没有受到儿童年龄(r= 0.03,p= 0.88)和儿童性别(t(30)=0.64,p= 0.53)的显著影响。
情绪调节与相干性之间的相关。接下来研究者考察了CoopP条件下的相干性与CH8和12中的CoopP与CompP,CoopP与CoopStr以及亲子的情绪调节之间的关系。在CH12中,控制孩子的性别,CoopP-CoopStr对比的相干性与儿童的情绪调节(p= 0.006,p= 0.004,p= 0.038,95%CI= [0.28,0.79])和父母重新评估(PR= 0.57,p= 0.004,padj.=0.038,95%CI= [0.24,0.79;图A.2c)均呈显著相关)。因此,在合作期间,与FPC中的成年陌生人相比,与父母更高的脑同步和更好的父母和孩子的情绪调节有关。相干性和情绪调节之间的其他相关没有达到显著水平。
中介分析。为了探究CH12脑信号相干性在CoopP-CoopStr条件比较、儿童情绪调节和父母重新评估之间三者之间的关系,研究者进行了中介分析,以检查相干性是否中介了父母重新评估与儿童情绪调节之间的关系(如图3)。儿童性别被纳入中介模型和因变量的协变量中。研究结果表明,当考虑CH12的相干性(CoopP-CoopStr对比)为中介变量时,父母重新评估对儿童情绪调节没有显著影响(B= 0.037,SE= 0.043,p= 0.39,95%CI[-0.051,0.126])。正如不包括零的偏差校正后的自举置信区间(B= 0.051,SE= 0.026,95%CI[0.011,0.115))所显示的那样,父母重新评估通过中介变量对儿童情绪调节的影响具有统计学意义。因此,在数学模型中,与陌生人儿童合作条件相比,使用重新评估策略的父母可以预测FPC在亲子合作条件下更高的脑同步,反过来这又对孩子的情绪调节产生积极影响。
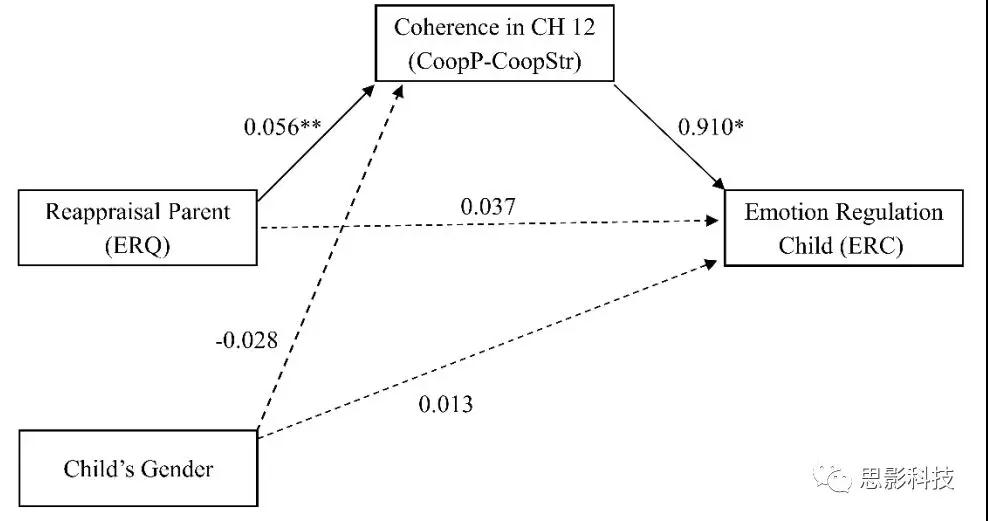
图3.父母重新评估与儿童情绪调节之间的关系是通过对比亲子合作与陌生成人儿童合作(CoopP-CoopStr)(N= 25)中的前极皮层中的相干性来中介的。实线和虚线分别代表显著和非显著效应。* p <0.05; ** p <0.01。ERC:情绪调节清单; ERQ:情绪调节问卷。
一句话总结:本研究表明,脑同步可能是一个支持在亲子互动过程中社交行为和情绪状态耦合以及儿童适应性情绪调节发展的神经机制基础。
参考文献:Vanessa Reindl,Brain-to-brainsynchrony in parent-child dyads and the relationship with emotion regulationrevealed by fNIRS-based hyperscanning
获取原文:关注“思影科技”公众号,回复“原文”或“培训”,获取原文pdf及补充材料下载链接,同时欢迎浏览我们的课程通知及数据处理业务介绍。(直接点击红色字即可):
第三届近红外脑功能数据处理培训班
第七届脑电数据处理基础班
更新通知:第二届脑电信号数据处理提高班
第一届功能磁共振提高班
思影数据处理业务四:EEG/ERP数据处理