≈Π‘Φ÷ίΝΔ¥σ―ßΨΪ…ώ≤Γ―ßœΒΫΧ ΎMeyersΒ»»Υ‘ΎMolecular Psychiatry‘”÷ΨΖΔ±μΖ«÷ό―ΣΆ≥Φ“Ήε÷–ΨΤΨΪ“άάΒΒΡΡΎ±μ–Ά―–ΨΩΓΘΩλΥΌbetaΫΎ¬…Θ®20-28HzΘ§“‘œ¬Φρ≥ΤbetaΘ©EEG’ώΒ¥ΜνΕ·Ω…Ρή «―–ΨΩ…ώΨ≠≥§–ΥΖή–‘(neuralhyperexcitability)Φ≤≤ΓΒΡ“≈¥Ϊ―ß”–”ΟΒΡΡΎ±μ–Ά(endophenotype)Θ§Αϋά®Έο÷ Ι”Ο’œΑ≠Θ®substance use disorders, SUDsΘ©ΓΘ»ΜΕχΘ§ΡΩ«ΑΜΙΈ¥Ε‘Ζ«“αΟάΙζ»Υ―ΣΆ≥Θ®African-American ancestry,AAΘ©ΒΡEEGΠ¬ΫΎ¬…ΒΡ“≈¥ΪΜυ¥ΓΫχ––―–ΨΩΓΘ“ρ¥ΥΘ§ΗΟ―–ΨΩΒΡΒΎ“ΜΡΩ±ξΘΚ¥”ΨΤΨΪ÷–ΕΨ“≈¥Ϊ―ßΚœΉς―–ΨΩΘ®The Collaborative Study on the Genetics of Alcoholism, COGAΘ©÷–Χτ―ΓAAΦ“ΆΞΘ§ΉωbetaΫΎ¬…Θ®ΖΔ…ζ”ΎΥΪ≤ύ’μ“ΕΘ§«Α»Υ―–ΨΩ÷–ΖΔœ÷¥ΥΫΎ¬…ΨΏ”–ΗΏΕ»ΒΡ“≈¥Ϊ–‘Θ©ΒΡ»ΪΜυ“ρΉιΙΊΝΣ―–ΨΩΘ®Genome-wideassociation study, GWASΘ©Θ§’β–©AAΦ“ΆΞ «ΉνΫϋ»ΖΕ®ΒΡ ήΒΫΨΤΨΪ“άάΒΘ®AlcoholDependence, ADΘ©ΚΆΙ≤Ά§ΖΔ…ζΒΡΆβΜ·’œΑ≠Θ®externalizingdisordersΘ§άΐ»γΘ§SUD,ADHDΘ©”ΑœλΒΡΦ“ΆΞ―υ±ΨΓΘΒΎΕΰΗωΡΩ±ξΘΚΧΫΥςGWASΖΔœ÷ΒΡΙΠΡήΚΆ––ΈΣ―ß“β“εΓΘ
»ΥάύΒΡΡ‘…ζάμ’ώΒ¥Ζ÷ΈΣ≤ΜΆ§ΒΡΤΒ¥χΘ®delta(1-3 Hz), theta (4-7 Hz), alpha (8-12 Hz), beta (13-28 Hz)ΚΆgamma(>29 Hz)Θ©Θ§ΟΩ“ΜΤΒ¥χΖ¥”≥ΝΥ≤ΜΆ§ΒΡ»ΪΡ‘Ή¥Χ§Θ®άΐ»γΘ§alphaΖ¥”≥ΝΥΖ≈Υ…Ή¥Χ§Θ§ΕχbetaΖ¥”≥ΝΥΨ·ΨθΉ¥Χ§Θ©ΓΘ”–―–ΨΩΖΔœ÷Θ§beta”κΦΗ÷÷ΆβΜ·’œΑ≠”–ΙΊΘ®»γΘ§AD,SUDs, ADHDΘ©ΓΘΦχ”Ύ¥Υ“‘ΦΑ“‘Άυ‘ΎΥΪ…ζΉ”―–ΨΩ÷–ΖΔœ÷ΒΡbetaΒΡΗΏΕ»“≈¥Ϊ–‘Θ§―–ΨΩ’Ώ»œΈΣΘ§beta «“ΜΗω”–”ΟΒΡΡΎ±μ–ΆΘ§”Ο”Ύ Ε±πΗωΧε’β–©’œΑ≠ΒΡ«±‘Ύ“≈¥Ϊ“ρΥΊΓΘ
÷°«Α”–―–ΨΩ±®ΗφΝΥΨΤΨΪ Ι”Ο’œΑ≠Θ®alcoholuse disordersΘ§AUDΘ©ΒΡbetaΘ®>19HzΘ©ΡήΝΩ¥φ‘Ύ≤ν“λΓΘ¥ΥΆβΘ§beta≤Μ ήΦ≤≤Γ―œ÷Ί≥ΧΕ»ΒΡ”ΑœλΓΘ”…”Ύ–οΨΤ’ΏΒΡΚσ¥ζ‘ΎΈΘœ’“ϊΨΤΩΣ Φ÷°«ΑΨΆ¥φ‘Ύ…ΐΗΏΒΡbetaΘ§“ρ¥ΥΘ§―–ΨΩ’ΏΦΌ…ηΘ§ΙΐΗΏΒΡbetaΙΠ¬ «œ»”ΎAUDΒΡΖΔ’ΙΘ§Εχ«“Θ§”κΖΔ’ΙΈΣAUDΒΡ«±‘Ύ“≈¥Ϊ«ψœρ «œύΙΊΒΡΘ§Φ¥betaΩ…Ρή «¥σΝΩ“ϊΨΤΒΡ‘≠“ρΘ§Εχ≤Μ «¥σΝΩ“ϊΨΤΒΡΫαΙϊΓΘBegleiterΚΆPorjesz»œΈΣbetaΩ…Ρή «¥σΡ‘ΤΛ≤ψ–ΥΖή-“÷÷Τ ßΚβΒΡΒγ…ζάμ÷Η±ξΘ§Εχ’β»ί“ΉΖΔ’ΙΈΣAUDΓΘΫχ“Μ≤Ϋ÷ß≥÷’β“ΜΦΌ…ηΒΡ «beta”κΤδΥϊ’œΑ≠ΒΡΙΊΝΣΘ§»γ––ΈΣ’œΑ≠Θ§ΕυΆ·ΓΔ«ύ…ΌΡξΒΡΆβΜ·’œΑ≠Θ§ADHDΘ§Άχ¬γ≥…ώΪΒ»ΓΘ
ΨΓΙήΥΪ…ζΉ”ΚΆΦ“Ήε―–ΨΩΖΔœ÷ΝΥbetaΒΡΗΏ“≈¥Ϊ–‘Θ®49-85%Θ©Θ§ΒΪœ ”–“≈¥Ϊ―ßΕ‘¥ΥΫχ––―–ΨΩΘ§ΡΩ«ΑΘ§Ϋω”–“ΜœνΖΔœ÷ΒΟΒΫ÷ΊΗ¥ΓΘ÷Ί“ΣΒΡ «Θ§Ε‘AA»ΚΧεΜΙΟΜ”–Ϋχ––EEGΒΡGWASΘ§“ρ¥Υ‘ΎAA»ΚΧε÷–ΜΙΟΜ”–ΚήΚΟΒΊΟη ωEEGΒΡœύΙΊ–‘ΧΊ’ςΒΡ“≈¥ΪΫαΙΙΓΘ‘ΎAA»ΚΧε÷–Ϋχ––“≈¥Ϊ―ß―–ΨΩ≥ΐΝΥΩΦ¬«ΒΫΙΪΙ≤Έά…ζ÷Ί“Σ–‘÷°ΆβΘ§ΜΙ“ρΈΣΦ≤≤ΓΒ»ΈΜΜυ“ρΤΒ¬ ΚΆΝ§Υχ≤ΜΤΫΚβΡΘ Ϋ¥φ‘ΎΗϋ¥σΒΡ“≈¥ΪΕύ―υ–‘ΚΆΫχΜ·≤ν“λ–‘ΓΘ¥ΥΆβΘ§AA“ϊΨΤ’Ώ“ΣΨ≠άζΗϋΕύ”κΨΤΨΪ”–ΙΊΒΡΈ ΧβΘ§Αϋά®…γΫΜΈ ΧβΘ§Φ≤≤ΓΚΆΥάΆωΘ§’βΥΒΟς–η“Σ Ε±πΦθ«αΒΦ÷¬Έ Χβ“ϊΨΤΖγœ’ΒΡ“ρΥΊΓΘ“ρΈΣ―–ΨΩΜυ±ΨΒΡ¥σΡ‘ΙΠΡή”κ»ΥάύΒΡ––ΈΣΚΆ’œΑ≠ΒΡΙΊœΒΒΡΉν÷’ΡΩ±ξ «ΧαΙ©Ε‘Υυ”–»ΥΒΡ‘ΛΖάΚΆ/ΜρΗ…‘ΛΘ§’β“ΜΈ ΧβΒΡάμ¬έ–η“ΣΦ”“‘ΫβΨωΓΘΦχ”ΎbetaΨΏ”–ΗΏΕ»Ω…“≈¥ΪΒΡ≤Δ«“”κΦΗ÷÷ΆβΜ·––ΈΣ“‘ΦΑSUDs”–ΙΊΘ§betaΒΡ“≈¥ΪΖ÷ΈωΩ…Ρή”–÷ζ”ΎΈ“Ο«άμΫβ”–“ΜœΒΝ–ΆβΜ·’œΑ≠Ζγœ’ΒΡΗωΧεΒΡΡ‘ΙΠΡήΓΘ“ρ¥ΥΘ§±Ψ―–ΨΩΒΡΒΎ“ΜΗωΡΩ±ξ «Ε‘AA»ΚΧεΫχ––EEGΒΡbetaΫΎ¬…ΒΡGWASΒΡ―–ΨΩΓΘΒΎΕΰΗωΡΩ±ξ «ΧΫΥςGWASΖΔœ÷ΒΡΙΠΡήΚΆ––ΈΣ―ß“β“εΓΘ
ΗΟ―–ΨΩ¥”COGA÷–―Γ‘ώ482ΉιΦ“ΆΞ÷–ΒΡ2382ΗωAAΉςΈΣ±Μ ‘Θ§Ε‘ΤδEEGΨ≤œΔΧ§ΒΡbetaΫΎ¬…Ϋχ––GWASΓΘΈΣΫχ“Μ≤ΫΟη ωΜυ“ρΖΔœ÷Θ§ΗΟ―–ΨΩΧΫΥςΝΥGWAS±δ“λΧεΒΡΙΠΡήΚΆΝΌ¥≤/––ΈΣ“β“εΓΘΫαΙϊœ‘ ΨΈΜ”Ύ»Ψ…ΪΧε3q26ΒΡΜυ“ρ«χΦδ…œΒΡ10ΗωœύΙΊΒΡΒΞΚΥή’ΥαΕύΧ§–‘Θ®SNPsΘ©Θ®r2> 0.9Θ©”κbetaΫΎ¬…œύΙΊΘ®P < 5 ΓΝ 10 - 8Θ©ΓΘΉνœ‘÷χœύΙΊΒΡSNP «rs11720449Θ®Π¬= -0.124Θ§p<4.5ΓΝ10-9Θ©Θ§“≤ «ΕΓθΘΒ®ΦνθΞΟΗΘ®Butyrylcholinesterase,BCHEΘ©‘Ύ«πΡ‘Ήι÷·÷–±μ¥οΒΡ ΐΝΩ–‘Ή¥ΈΜΒψΓΘ10÷÷÷–ΒΡ4÷÷»ΪΜυ“ρΉιSNPs”κCOGAΒΡAAΦ“ΆΞΒΡΨΪ…ώ’œΑ≠ΨΤΨΪ“άάΒ’οΕœΚΆΆ≥ΦΤΫαΙϊ”–ΙΊΝΣΘ§Τδ÷–ΝΫΗωΘ®rs13093097Θ§rs724372Θ©“―‘ΎΕάΝΔΒΡAA±Μ ‘÷–±Μ÷ΊΗ¥ΓΘ‘ΎAA«ύ…ΌΡξΘ®ά¥Ή‘COGAΦ“ΆΞΒΡΚσ¥ζΘ©ΒΡΖ÷Έω÷–±μΟςΘ§rs11720469”κ÷ΊΕ»“ϊΨΤΘ®ΤΒ¬ ΈΣ24hΡΎ“ϊ”Ο5ΤΩΨΤ“‘…œΘ©”–ΙΊΓΘΗΟ―–ΨΩΈΣ3q26÷–ΒΡΜυ“ρ±δ“λ‘Ύ…ώΨ≠ΚΆ––ΈΣ“÷÷Τ÷–ΒΡΉς”ΟΧαΙ©ΝΥ÷ß≥÷ΓΘ’β–©–¬ΒΡΜυ“ρΖΔœ÷«ΩΒςΝΥ‘ΎΜυ“ρ―–ΨΩ÷–Φ”»κAA÷÷»ΚΒΡ÷Ί“Σ–‘“‘ΦΑΡΎ±μ–ΆΖΫΖ®ΒΡ–ß”ΟΘ§“‘‘ω«ΩΈ“Ο«Ε‘≥…ώΪ“ΉΗ––‘Μζ÷ΤΒΡΝΥΫβΓΘ
Β―ιΖΫΖ®ΘΚ
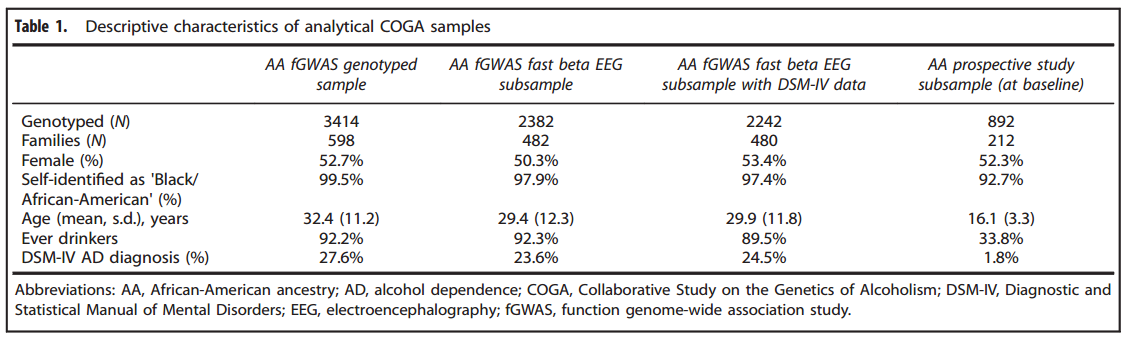
ΙΊ”ΎΨΤΨΪ÷–ΕΨ“≈¥Ϊ―ßΒΡΚœΉς―–ΨΩ
≤Έ”κ’ΏΫ” ήΨΤΨΪ÷–ΕΨ“≈¥Ϊ―ßΒΡΑκΫαΙΙΜ·ΤάΙάΘ®TheSemi-Structured Assessment for the Genetics of Alcoholism, SSAGAΘ©Θ§’β «“ΜΗωΕύ’οΕœ–‘ΒΡΖΟΧΗΓΘΡξΝδ‘Ύ18Υξ“‘œ¬ΒΡΗωΧεΫ” ή«ύ…ΌΡξΑφΒΡSSAGAΓΘΜΙ“Σ ’Φ·ΥϊΟ«ΒΡDNAΚΆEEGΓΘ¥”GWAS ΐΨί÷–Χα»ΓΒΡ÷ς≥…Ζ÷Θ®Principalcomponents, PCsΘ©±Μ”Οά¥‘Ύ»ΪΜυ“ρΖ÷–Ά―υ±Ψ÷–Ζ÷ΉιΈΣ≤ΜΆ§ΒΡΉφœ»―ΣΆ≥Θ§Ά§ ±“≤ «―Γ‘ώAAΦ“ΆΞΒΡΜυ¥ΓΓΘΖ÷ΈωΒΡ―υ±ΨΑϋά®ά¥Ή‘482ΗωΦ“ΆΞΒΡ2382ΗωΗωΧεΘ®Table1Θ©Θ§”–99.5%ΒΡAAΗωΧεΘ®Ά®ΙΐPCsΕ®“εΘ©Ή‘Έ“Ε®“εΈΣΚΎ»Υ/Ζ«“αΟάΙζ»ΥΘ§Τδ÷–27.6%ΖϊΚœAD±ξΉΦΘ§ΤδΥϊΙ≤Ά§ΖΔ…ζΒΡ“©Έο Ι”ΟΚΆΆβΜ·’œΑ≠“≤ «÷Ί“ΣΒΡ±ξΉΦΘ§άΐ»γΘ§Ω…Ω®“ρ“άάΒΘ§ADHDΓΘ
2.EEGΦ«¬Φ
Φ«¬Φ±’―έΨ≤œΔEEG4.25 minΘ§Ζ÷Έω256sΒΡΝ§–χ ±ΦδΕΈΓΘ±Ψ―–ΨΩ÷ς“ΣΙΊΉΔΒΡ «Ζ÷≤Φ‘ΎΥΪ≤ύ’μ“Ε…œΘ®O1-O2Θ©Θ§Ψ≠ΙΐΕ‘ ΐΉΣΜΜΒΡΨχΕ‘ΒΡbetaΫΎ¬…Θ®20-28HzΘ©ΒΡΙΠ¬ ΓΘ
3. Μυ“ρ–ΆΘ§Ιι“ρΚΆ÷ ΝΩΩΊ÷Τ
Ι”ΟIllumina2.5M’σΝ–Θ®Illlumina,San Diego, CA, USAΘ©‘Ύ“≈¥ΪΦ≤≤Γ―–ΨΩ÷––ΡΕ‘598ΗωΦ“ΆΞΒΡ3414Ηω»ΥΫχ––Μυ“ρ–ΆΖ÷άύΓΘΜυ“ρΖ÷–Ά¬ –Γ”Ύ98%ΜρΈΞΖ¥ Hardy-WeinbergΤΫΚβΘ®p< 10-6Θ©Μρ¥ΈΒ»ΈΜΜυ“ρΤΒ¬ Θ®minorallele frequencyΘ§MAFΘ©<3ΒΡSNPs±Μ≈≈≥ΐΓΘ≤ΜΖϊΚœΟœΒ¬ΕϊΘ®MendelianΘ©“≈¥ΪΕ®¬…ΒΡ ΐΨί±Μ“Τ≥ΐΘ§ Ι”ΟSHAPEITΚΆIMPUTE2Θ§ ΐΨί±ΜΙι“ρΒΫ1000ΗωΜυ“ρΉιΘ®hg19Θ©ΓΘΫ”œ¬ά¥Θ§Μυ“ρ–ΆΗ≈¬ Γί 0.9±ΜΗΡ±δΈΣΜυ“ρ–ΆΓΘΗυΨίWetherillΒ»»ΥΒΡΟη ωΘ§‘Ύ±ΜΖ÷άύΒΡSNPs÷–Θ§Mendelian¥μΈσ±Μ÷Ί–¬ΙάΦΤΚΆΫβΨωΓΘΙι“ρ–≈œΔΒΟΖ÷ < 0.3ΜρMAF < 0.03ΒΡSNPs‘ΎΥφΚσΒΡΖ÷Έω÷–±Μ…Ψ≥ΐΓΘ
4.GWAS
Ι”ΟœΏ–‘ΜλΚœΡΘ–ΆΕ‘‘ΎGWAFΘ®Genome-wideassociation analyses with familyΘ©÷–ΒΡ12972748ΗωSNPsΫχ––GWASΖ÷ΈωΘ§≤Δ«“Ρ…»κ“≈¥ΪΙΊΝΣΨΊ’σΘ®geneticrelatedness matrixΘ©ά¥ΩΊ÷ΤΦ“ΆΞ―υ±Ψ÷–ΒΡ«Ή‘ΒΙΊœΒΓΘ“ρΈΣ–‘±πΚΆΨ≠ΙΐΕ‘ ΐΉΣΜΜΒΡΡξΝδΘ®EEGΦ«¬Φ ±Θ©ΕΦ”κΠ¬ΫΎ¬…”–ΙΊΘ®p< 0.001Θ©Θ§“ρ¥Υ‘ΎΡΘ–Ά÷–Θ§ΥϊΟ«ΉςΈΣ–≠±δΝΩ¥φ‘ΎΓΘΈΣΫχ“Μ≤ΫΦθ…Ό”…”Ύ»ΚΧεΖ÷≤ψΒΦ÷¬ΒΡΦΌ―τ–‘Θ§¥”SNPRelateΦΤΥψΒΟΒΫΒΡ«Α10ΗωPCsΘ®PC1-PC10Θ©“≤ΉςΈΣΡΘ–Ά–≠±δΝΩΘ§ΦΌ…ηΝΥ“ΜΗωΕνΆβΒΡ“≈¥ΪΡΘ–ΆΓΘΉνΚσ Ι”Οp< 5ΓΝ10-8±μ ΨΈΣ»ΪΜυ“ρΉιœ‘÷χ–‘ΒΡψ–œόΓΘάϊ”Ο»ΪΜυ“ρΉιΗ¥‘”–‘ΧΊ’ςΖ÷ΈωΘ®Genome-widecomplex trait analysis, GCTAΘ©»ΖΕ®―υ±Ψ÷–Π¬ΫΎ¬…ΒΡSNP“≈¥Ϊ¬ ΓΘΫΪ“≈¥ΪΙΊΝΣΨΊ’σΡ…»κΦ“ΉεΨέάύΒΡ“≈¥ΪΙάΦΤ÷–ΓΘ
5. ΙΠΡήΖ÷Έω
‘Ύ”ΔΙζΡ‘±μ¥ο–≠ΜαΘ®UKBrain Expression ConsortiumΘ§BRAINEACΘ©÷–Θ§Έ“Ο«Φλ―ιΝΥ”κbetaΫΎ¬…Ήνœ‘÷χΙΊΝΣΒΡ±δ“λΧε «Ζώ «“ΜΗω±μ¥οΒΡ ΐΝΩ–‘Ή¥Μυ“ρΉυΘ®expressionquantitative trait locusΘ§eQTLΘ©ΓΘBraineac ’Φ·ΝΥά¥Ή‘134Ηω≈Ζ÷ό―ΣΆ≥ΒΡ…ώΨ≠≤Γάμ―ß’ΐ≥ΘΗωΧεΒΡ ΐΨίΘ®Table3Θ©Θ§≤ΔΕ‘10Ηω≤ΜΆ§ΒΡΡ‘«χΫχ––ΤάΙάΓΘ÷Μ”–”κΠ¬ΫΎ¬…ΉνœύΙΊΒΡΒΞΗωΒΡSNP≤≈‘ΎBraineac±ΜΖ÷Έω¥”Εχ ΙΒΟΕύ÷Ί≤βΝΩ¥οΒΫΉν–ΓΜ·Θ§Υυ”–ΒΡp÷ΒΕΦΨ≠ΙΐBonferonniΫΟ’ΐΓΘ‘ΎGenotype-TissueExpression ProjectΘ®GTExΘ© ΐΨίΩβ÷–Θ§Έ“Ο«÷ΜΖ÷ΈωΝΥΨ≠ ήΉΓΕύ¥Έ≤βΝΩΜΙ¥οΒΫœ‘÷χΥ°ΤΫΒΡ“≈¥Ϊ-Π¬ΙΊΝΣΘ§“‘»ΖΕ®eQTLΒΡΖΔœ÷ΓΘ
6. “ϊΨΤ––ΈΣ
Έ“Ο«»ΖΕ®ΝΥ”κΠ¬œύΙΊΒΡ»ΪΜυ“ρΉι±δ“λ «Ζώ“≤”κ―υ±ΨΘ®ά¥Ή‘480ΗωΦ“ΆΞ÷–ΒΡ2242»ΥΘ©÷–ΒΡDSM-IV ADΘ®Diagnosticand Statistical Manual of Mental DisordersΘ©œύΙΊΓΘ Ι”ΟSAS9.4»μΦΰΫχ–– ΐΨίΖ÷ΈωΓΘΗυΨίΡξΝδΘ§–‘±πΘ§«Ή‘ΒΙΊœΒΚΆPC1-PC10Ε‘¬ΏΦ≠ΜΊΙιΡΘ–ΆΫχ––Βς’ϊΓΘΈ“Ο«Φλ―ιΝΥΥυ”–ΒΡΠ¬ΒΡ”κ»ΪΜυ“ρΉιœ‘÷χΒΡSNPsΒΡœύΙΊΓΘ Ι”ΟPnorm≥Χ–ρΕ‘ΗωΧεΒΡp÷ΒΫχ––Βς’ϊΘ§’β“Μ≥Χ–ρΑϋά®SNPsΒΡLDΫαΙΙΘ®Linkagedisequilibrium patternsΘ©ΚΆ«Ή τΒΡ≥ι―υΓΘ
Ϋ”œ¬ά¥Θ§‘Ύ≤ΜœύΙΊΗωΧεΒΡΕάΝΔ―υ±Ψ÷–Θ®±Μ’–ΡΦά¥―–ΨΩΑΔΤ§άύΓΔΩ…Ω®“ρΜρADΒΡΜυ“ρΘ§ ΐΨίΩ…‘ΎdbGaP…œΙΪΩΣΖΟΈ Θ©Θ§Έ“Ο«―–ΨΩGWAS±δ“λ «Ζώ”κDSM-IVADœύΙΊΝΣΓΘ÷Μ”–Ω…“‘ΜώΒΟAD ΐΨίΒΡAAΗωΧε≤≈±ΜΡ…»κΖ÷ΈωΘ®1346ΗωADΚΆ461ΗωΩΊ÷ΤΉι±Μ ‘Θ©ΓΘ
ΉνΚσΘ§ Β―ι“Σ«σ≤Έ”κ’ΏΘΚΓΑΥΦΩΦΙΐ»ΞΒΡ12Ηω‘¬Θ§”–Εύ…Ό¥Έ «‘Ύ24–Γ ±ΡΎΚ»5ΤΩΜρ’Ώ5ΤΩ“‘…œΒΡΨΤΘΩΓ±Θ®ΑΉΒΡ¬πΘΩΘ©Θ§―Γœν¥”ΓΑ¥”≤ΜΓ±ΒΫΓΑΟΩΧλΓ±ΓΘ‘Ύ212ΗωΦ“ΆΞ÷–892»Υ÷–Θ§”–33.8%ΒΡ»Υ‘χΨ≠Κ»Ιΐ¥σΝΩΒΡΨΤΓΘ Θ”ύΒΡ66.2% «0ΓΘ”…”Ύ―υ±ΨΝΩœύΕ‘Ϋœ…ΌΘ§÷ΜΖ÷Έω”κΠ¬ΉνœύΙΊΒΡSNP“‘Ήν–ΓΜ·Εύ÷ΊΦλ―ιΓΘ‘ΎMplus7.4÷–Θ§ Ι”ΟΕ‘ ΐΉΣΜΜΚσΒΡ÷ΊΕ»“ϊΨΤ’ΏΕ‘¥ΥΙΊΝΣΫχ––Φλ―ιΘ§≤ΔΗυΨί«Ή‘ΒΙΊœΒΓΔ–‘±πΓΔΡξΝδΚΆPC1-PC10Ε‘ΡΘ–ΆΫχ––Βς’ϊΓΘ
7. ¬ΚσΦλ―ι
œ»«Α―–ΨΩΖΔœ÷BCHEΘ®Μρ÷ή±Ώ«χ”ρ:3q26Θ©”κbeta“‘ΦΑAUDΘ®Αϋά®ADHDΘ§Ω…Ω®“ρ Ι”ΟΈ ΧβΘ©ΒΡ––ΈΣ―ßœύΙΊΝΣΓΘΈΣΝΥ»ΖΕ®3÷÷’œΑ≠Θ®DSM-IV,DSM-IV ADHD, DSM-IV CoDΘ©ΗςΉ‘ «ΖώΩ…“‘Ϋβ Άœ‘÷χΒΡbeta–≈Κ≈Θ§Έ“Ο«Ζ÷±πΕ‘3÷÷Φ≤≤ΓΒΡbetaΒΡGWASΫχ––ΝΥ ¬ΚσΦλ―ιΘ§Τδ÷–Θ§ΟΩ÷÷’œΑ≠ΉςΈΣΡΘ–ΆΒΡ–≠±δΝΩΓΘ
ΫαΙϊΘΚ
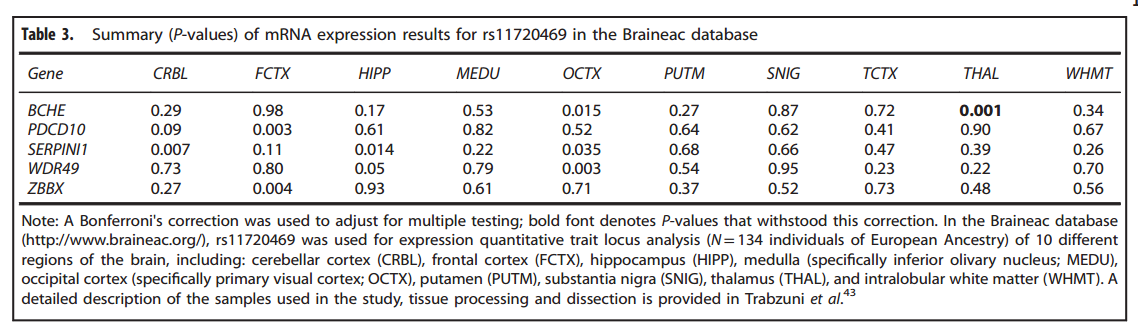
betaΫΎ¬…ΒΡGWAS
ΈΜ”Ύ3q26…œΒΡΜυ“ρΦδΗτ«χ”ρΘ®Chr3, 166 471 942®C166 489 551Θ©ΒΡ10ΗωSNPsΘ®Μυ”ΎΖ«÷ό―ΣΆ≥÷–ΒΡhg191000Μυ“ρΉιΒΡΥυ”–10ΗωSNPs≥…Ε‘œύΙΊr2> 0.9Θ©”κbetaΫΎ¬…œύΙΊΘ®p< 5ΓΝ10-8Θ©Θ®Table2Θ§Figure 1Θ§Figure 2Θ©ΓΘΉνœ‘÷χΒΡSNP «rs11720469Θ®p< 4.5ΓΝ10-9Θ©Θ§¥Έ“ΣΒ»ΈΜΜυ“ρΘ®GΘ©”κbeta≥ ΗΚœύΙΊΘ®Π¬= -0.124Θ©ΓΘΆΦ2’Ι ΨΝΥGWAS–≈Κ≈“‘ΦΑ“―÷ΣΜυ“ρ…œΓΔœ¬”ΈΒΡ–≈Κ≈Θ§Αϋά®BCHE,PDCD10, WDR49, SERPINI1, SERPINI2ΚΆZBBXΓΘGABRA2“≤”κbetaΫΎ¬…œύΙΊΘ§ΒΪ≤Μ «»ΪΜυ“ρΉιΥ°ΤΫ…œΘ®p< 0.01Θ©ΓΘGCTAΖΔœ÷Θ§‘ΎbetaΫΎ¬…÷–Θ§”–33.8%ΒΡ±δ“λΘ®s.e.:0.014, p < 5.5ΓΝ10-17Θ© «”…”Ύ»ΪΜυ“ρΉιΥ°ΤΫΒΡSNPsΘ®œΝ“ε“≈¥Ϊ¬ Θ©‘λ≥…ΒΡΓΘ

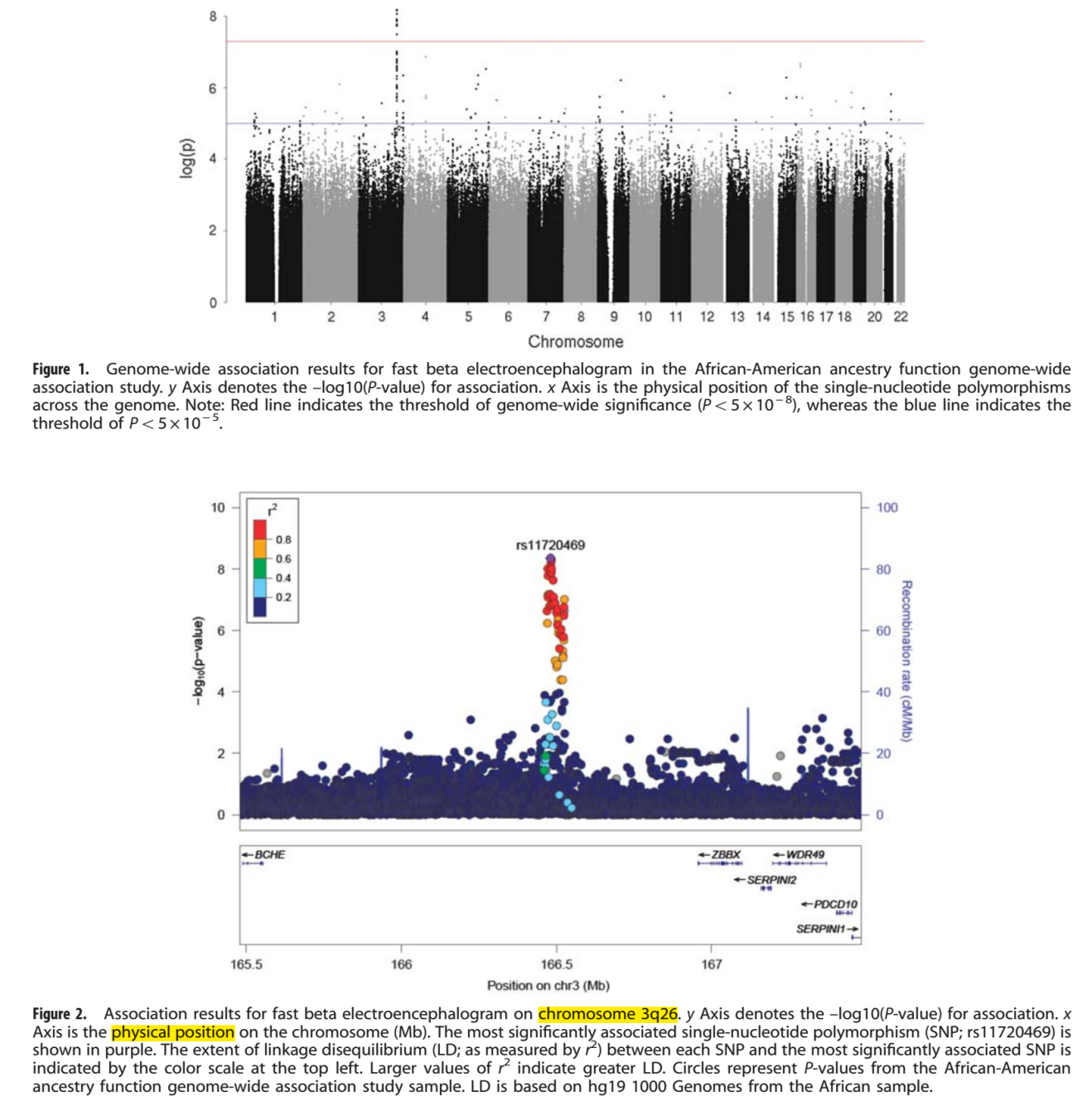
2. ΙΠΡήΖ÷Έω
‘ΎBraineac ΐΨίΩβ÷–Θ§rs11720469”κBCHE,PDCD10, SERPINI1, WDR49ΚΆZBBXΒΡmRNAΒΡ±μ¥οœύΙΊΓΘΤδ÷–Θ§Ψ≠ΙΐBonferroniΫΟ’ΐ“‘ΚσΘ§÷Μ”–“ΜΗωΫαΙϊ“άΨ…œ‘÷χΘΚrs11720469 ««πΡ‘Ήι÷·÷–BCHE±μ¥οΒΡeQTLΘ®p= 4.20ΓΝ10-4Θ©ΓΘ¥Έ“ΣΒ»ΈΜΜυ“ρ”κΦθ…ΌΒΡmRNA±μ¥ο”–ΙΊΘ®Table3Θ©ΓΘ‘ΎGTEx ΐΨίΩβ÷–Θ§rs11720469”κΡ‘Ήι÷·÷–BCHEΒΡ±μ¥ο”–ΙΊΘΚ¥σΡ‘ΤΛ≤ψΘ®p< 0.007Θ©Θ§Έ≤Ή¥ΚΥΘ®p< 0.005Θ©ΓΘHaploRegV4.1÷Η≥ω‘ΎROADMAP±μΙέΜυ“ρΉι―ß ΐΨί÷–Θ§rs11720469ΗΡ±δΝΥ“Μ–©œΗΑϊΒΡΒςΩΊΡΘ ΫΓΘ
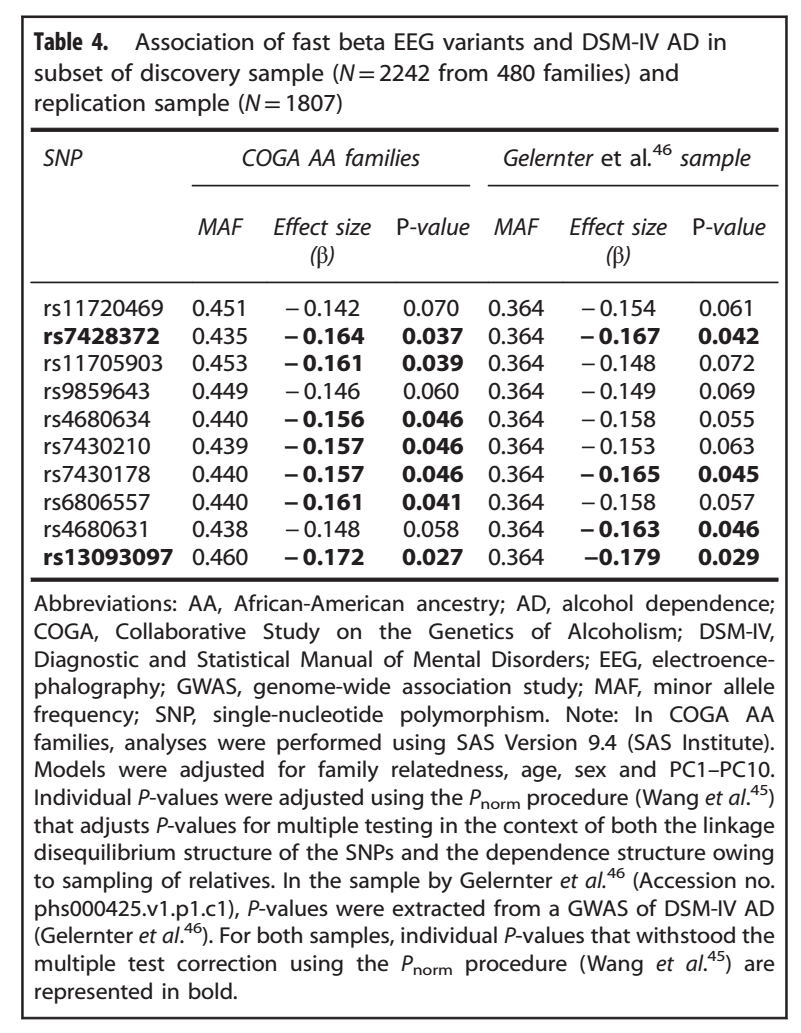
3.“ϊΨΤ––ΈΣ
‘ΎGWASΒΡΉ”―υ±Ψ÷–Θ§”κbetaΫΎ¬…œύΙΊΒΡ10Ηω±δ“λ÷–ΒΡ7Ηω“≤”κAD”–ΙΊΘ®Table4Θ©ΘΜ¥Έ“ΣΒ»ΈΜΜυ“ρ”κΦθ…ΌΒΡADΖγœ’”–ΙΊΘ§±μ Ψ“ΜΕ®ΒΡ±ΘΜΛΉς”ΟΓΘΩΦ¬«ΒΫ‘Ύ’β10ΗωSNPs÷–Ιέ≤λΒΫΒΡΗΏLDΘ§’β–©p÷ΒΗυΨί Ι”ΟPnorm≥Χ–ρΙάΦΤΒΡ≤β ‘ ΐΝΩΘ®1.2Θ©Ϋχ––Βς’ϊΘ§Τδ÷–4Ηω±δ“λΨ≠ΙΐΕύ÷ΊΫΟ’ΐΚσ≤ν“λ“ά»Μ¥φ‘ΎΘΚrs7428372Θ®Π¬ΘΚ-0.164Θ§p < 0.037Θ©Θ§rs11705903Θ®Π¬ΘΚ-0.161Θ§p< 0.039Θ©Θ§rs6806557Θ®Π¬ΘΚ-0.161Θ§p< 0.041Θ©Θ§rs13093097Θ®Π¬ΘΚ-0.172Θ§p< 0.027Θ©ΓΘ
‘ΎΕάΝΔ―υ±Ψ÷–Θ§10ΗωSNPs÷–ΒΡ4Ηω”κAD”–ΙΊΘ®Table4Θ©ΓΘΨ≠ΙΐΕύ÷Ί±»ΫœΫΟ’ΐΚσΘ§Τδ÷–2ΗωSNPsΒΡœύΙΊ“ά»Μ¥φ‘ΎΘΚrs7428372Θ®Π¬ΘΚ-0.167Θ§p< 0.042Θ©Θ§rs13093097Θ®Π¬ΘΚ-0.179Θ§p< 0.029Θ©Θ§¥Έ“ΣΒ»ΈΜΜυ“ρ”κΦθ…ΌΒΡADΖγœ’”–ΙΊΘ§±μ ΨΉ≈±ΘΜΛΉς”ΟΓΘ
ΉνΚσΘ§ά¥Ή‘COGAΦ“ΆΞΒΡ«ύ…ΌΡξ/Ρξ«α≥…»ΥΒΡAAΉ”―υ±ΨΘ§”–“ΜΗωΜρΕύΗω¥Έ“ΣΒ»ΈΜΜυ“ρΒΡΗ±±Ψ”κ÷ΊΕ»“ϊΨΤΒΡΦθ…Ό”–ΙΊΘ®Π¬ΘΚ-0.064Θ§p < 0.035Θ©Θ§ΒΪ «–ß”ΠΝΩ≤Μ¥σΓΘ
4. ¬Κσ±»ΫœΖ÷Έω
‘Ύ÷ς“ΣΒΡGWAS―υ±Ψ÷–Θ§betaΡήΝΩ‘ΫΗΏΒΡΗωΧε‘ΫΩ…ΡήΖϊΚœDSM-IVAD, CoDΚΆADHD±ξΉΦΘ®all p < 0.001Θ§BonferroniΫΟ’ΐΘ©ΓΘΈΣ»ΖΕ®Τδ÷–“Μ÷÷Φ≤≤Γ «ΖώΩ…“‘Ϋβ Άbeta÷–Ιέ≤βΒΫΒΡœ‘÷χ–≈Κ≈Θ§Έ“Ο«Ζ÷±πΫχ––ΝΥ3÷÷’œΑ≠ΒΡbetaΫΎ¬…ΒΡGWASΒΡ ¬ΚσΦλ―ιΘ§ΟΩ“ΜΦλ―ιΕΦΫΪ’β–©Μυ“ρ–Ά÷°“ΜΉςΈΣ–≠±δΝΩΓΘADΉςΈΣ–≠±δΝΩΘ§3q26”κbetaΒΡœύΙΊ»‘¥φ‘ΎΘ§ΒΪ”––©ΫΒΒΆΘ®rs11720469ΘΚΠ¬ΘΚ-0.12Θ§pΘΚ2.2ΓΝ10-8Θ©ΓΘDSM-IVCoDΉςΈΣ–≠±δΝΩΘ§œύΙΊ–‘»‘‘ΎΘ§ΒΪ «”–«αΈΔΦθ…ΌΘ®rs11720469ΘΚΠ¬ΘΚ-0.122Θ§pΘΚ1.3*10-8Θ©ΓΘDSM-IV ADHDΉςΈΣ–≠±δΝΩΘ§œύΙΊ–‘Φθ…ΌΘ®rs11720469ΘΚΠ¬ΘΚ-0.088Θ§pΘΚ6*10-4Θ©ΓΘ
Ϋα¬έΘΚ
’βœν―–ΨΩ «‘ΎAA―υ±Ψ÷–Ϋχ––ΒΡΘ§ΈΜ”Ύ3q26ΒΡΜυ“ρΦδΗτ«χΦδ…œΒΡ–≈Κ≈”κbetaΫΎ¬…¥φ‘ΎΙΊΝΣΓΘΉνœ‘÷χΒΡSNP «“Μ÷÷BCHEΒΡeQTLΘ§Υϋ «“Μ÷÷”κ“÷÷Τ’œΑ≠”–ΙΊ≤Δ‘Ύ«πΡ‘÷–±μ¥οΒΡΜυ“ρΘ§«πΡ‘ «¥σΡ‘÷–betaΫΎ¬…ΚΆAUDΒΡΚΥ–ΡΡ‘«χΓΘ¥ΥΆβΘ§‘Ύ“―Ψ≠ΖΔœ÷ΒΡ―υ±ΨΚΆΕάΝΔ―υ±Ψ÷–Θ§betaΫΎ¬…»ΪΜυ“ρΉιœύΙΊΒΡ±δ“λΘ®rs7428372ΚΆrs13093097Θ©”κAD”–ΙΊΝΣΓΘ’β–©÷ΛΨίΈΣ3q26ΡΎΒΡ“≈¥Ϊ±δ“λ‘Ύ…ώΨ≠≥§–ΥΖή–‘ΚΆ“‘≥εΕ·–‘ΈΣΧΊ’ςΒΡΦ≤≤Γ÷–ΥυΤπΒΡΉς”ΟΧαΙ©ΝΥ÷ß≥÷ΓΘΝμΆβΘ§±Ψ―–ΨΩ÷ΛΟςΝΥΡΎ±μ–ΆΖΫΖ®ΒΡΉς”ΟΘ§ΕχbetaΫΎ¬…ΒΡ“≈¥Ϊ―ßΖΔœ÷ΧαΙ©ΝΥ“ΜΗω«±‘ΎΒΡ…ζΈο―ßΦΌ…ηΘ®Φ¥…ώΨ≠≥§–ΥΖή–‘Θ©Θ§ΥϋΩ…“‘‘ω«ΩΈ“Ο«Ε‘ΙΠΡή–‘¥σΡ‘ΜΊ¬ΖΚΆAUD“‘ΦΑœύΙΊ––ΈΣΒΡ“ΉΗ––‘Μζ÷ΤΒΡάμΫβΓΘ
Έ¥ά¥―–ΨΩΖΫœρΘΚ
1Θ°ΗΟ―–ΨΩ÷–ΒΡΖΔœ÷±Ί–κ‘ΎΗϋ¥σ―υ±ΨΝΩΒΡAAΗωΧε÷–Ϋχ––÷ΊΗ¥ΓΘΈ¥ά¥ΒΡ―–ΨΩ”ΠΗΟΩΦ≤λΜυ“ρ±δ“λΕ‘ΖΔ”ΐΙΐ≥Χ÷–betaΫΎ¬…ΙλΦΘΒΡ”ΑœλΘ§“‘»ΖΕ®ΧΊΕ®ΡξΝδΒΡ”Αœλ“‘ΦΑ’β–©”ΑœλΚΆ/ΜρΨΪ…ώ≤ΓάμΩΣ ΦΘ®AUDΓΔADHDΓΔCoDΘ©÷°ΦδΒΡΝΣœΒΓΘ
2Θ°Έ¥ά¥ΒΡΉίœρ―–ΨΩΩ…“‘ΧΫΥςADHDΓΔ3q26ΚΆbetaΫΎ¬…÷°ΦδΒΡΝΣœΒΘ®άΐ»γΘ§betaΫΎ¬…Ω…“‘ΒςΫΎ3q26ΚΆADHD÷°ΦδΒΡΝΣœΒ¬πΘΩΘ©
3Θ°ΥφΉ≈betaΫΎ¬…”κΡω≥ίΕ·ΈοΚΆ»ΥάύΒΡCoDΝΣœΒ‘Ύ“ΜΤπΘ§”ΠΗΟΫχ“Μ≤ΫΧΫΥςBCHEΓΔbetaΫΎ¬…ΚΆCoD÷°ΦδΒΡΟςœ‘ΝΣœΒΓΘ
Meyers, J. L., Zhang, J., Wang, J. C., Su, J., Kuo, S. I.,Kapoor, M., ... & Kamarajan, C. (2017). An endophenotype approach to thegenetics of alcohol dependence: a genome wide association study of fast betaEEG in families of African ancestry. Molecularpsychiatry, 22(12),1767.
Doi: https://doi.org/10.1038/mp.2016.239

ΜΕ”≠ΈΔ–≈…®ΟηΕΰΈ§¬κΙΊΉΔΈ“Ο«
Μώ»Γ‘≠ΈΡΘΚΙΊΉΔΓΑΥΦ”ΑΩΤΦΦΓ±ΙΪ÷ΎΚ≈Θ§ΜΊΗ¥ΓΑ‘≠ΈΡΓ±ΜρΓΑ≈ύ―ΒΓ±Θ§Μώ»Γ‘≠ΈΡpdfΦΑ≤Ι≥δ≤ΡΝœœ¬‘ΊΝ¥Ϋ”Θ§Ά§ ±ΜΕ”≠δ·άάΈ“Ο«ΒΡ≈ύ―ΒΆ®÷Σ“‘ΦΑ ΐΨί¥Πάμ“ΒΈώΫι…ήΘ®ΒψΜςœ¬ΖΫ≥»…ΪΉ÷Φ¥Ω…δ·άάΘ©:
ΒΎΨ≈ΫλΡ‘Βγ ΐΨί¥ΠάμΜυ¥ΓΑύ
ΒΎ»ΐΫλΡ‘Βγ–≈Κ≈ ΐΨί¥ΠάμΧαΗΏΑύ
ΒΎΥΡΫλΫϋΚλΆβΡ‘ΙΠΡή ΐΨί¥ΠάμΑύ
ΒΎ“ΜΫλ―έΕ· ΐΨί¥ΠάμΑύ
ΒΎ °ΫλΙΠΡή¥≈Ι≤’ώ ΐΨί¥ΠάμΜυ¥ΓΑύ
ΒΎΝυΫλ¥≈Ι≤’ώΡ‘Άχ¬γ ΐΨί¥ΠάμΑύ
ΒΎΕΰΫλΕ·Έο¥≈Ι≤’ώΡ‘”Αœώ ΐΨί¥ΠάμΑύ
ΒΎΥΡΫλ¥≈Ι≤’ώΡ‘”ΑœώΫαΙΙΑύ
ΒΎΈεΫλ¥≈Ι≤’ώΟ÷…Δ’≈ΝΩ≥…œώ ΐΨί¥ΠάμΑύ
ΥΦ”Α ΐΨί¥Πάμ“ΒΈώΥΡΘΚEEG/ERP ΐΨί¥Πάμ