摘要:社会支配地位是社会等级制度的主要组织原则,有利于支配个体优先获得资源。在所有群体中,如果个体在社会情境中首先行动,并快速行动可以提供进化优势,那么它们更有可能成为支配者。瑞士洛桑联邦技术研究所生命科学学院脑科学研究所心理物理学实验室和行为遗传学实验室Carmen Sandi和Janirda Cruz等人于2018年8月在Cerebral Cortex(IF:6.308)发表文章。该文章主要研究快速决策行为(fastdecision-making)是否是支配者在社会情境之外的行为倾向。在了解了被试的社会支配动机表征之后,实验发现在各种决策任务中,社会支配地位较高的男性在不丧失准确性的情况下,反应速度都快于低支配地位的男性;但在简单反应任务中没有差异。研究继而选择一个决策任务,并应用高密度EEG通过事件相关电位来评估大脑活动的时间动态性。结果发现在决策任务中,高支配型个体的快速反应与刺激后约240 ms的显著激活脑信号有关。源定位分析显示高支配型个体在左侧岛叶和扣带回、右颞下角回和右角回(left insula and in the cingulate, right inferior temporal and rightangular gyri)的激活高于低支配型个体。我们的研究结果表明无论社会背景如何,在决策任务中迅速作出反应是社会倾向的一个生物指标。
关键词:高密度脑电图、领导能力、反应时、社会等级(Social hierarchy)
引言
社会等级制度在社会物种中无处不在。有研究显示,在竞争环境中,高主导地位的人在决策方面比低主导地位的人更快。为了验证人类的主导特征与行动的迅速性有关的假设,我们首先通过普遍使用的PersonalityResearch Form dominance subscale问卷(PRF-d)来描述个体的社会主导动机。假设是,在对涉及认知挑战的任务做出决定时,高主导地位的人会有更短的反应时间,而在执行简单的反应任务(simplereaction task, SRT)时,高与低主导个体没有差异。考虑到众所周知的性别差异,我们关注男性。因此,我们进行了一系列的实验,以评估人们在支配动机中得分高或低的情况是否会在不同的决策过程和SRT中做出不同反应。最后,我们选择的实验条件以揭示个体在执行与高密度脑电图(EEG)研究相关的事件相关电位(ERP)研究中存在反应时间的组间差异,以分析大脑激活的时间动态的潜在差异。
方法
被试:研究者从洛桑瑞士联邦理工学院(EPFL)和洛桑大学(UNIL)招募240名男性被者,并完成五项实验中的其中一项。根据他们在PRF-d(参见人格测验)中的得分是否低于或高于9 分(从412名学生问卷中获得的PRF-d中值),被试被分为低支配型或高支配型。所有被试身体健康状况良好,没有内科疾病和神经或精神病史。此外,被试测试视觉优势,并完成了标准化利手问卷。通过双眼的Freiburg视力测试测量,我们验证所有被试都有至少1.0的良好视力。
人格测验:实验前几天被试填写人格研究量表支配性子量表[PRF-d(Jackson1974)的在线问卷(www.qualtrics.com)评估其社会支配动机。由于特质焦虑与社会支配性相关并能预测行为结果,因此我们要求被试填写Speilberger的状态-特质焦虑问卷(STAI),并在统计分析中评估了STAI-T以控制这个变量。
行为实验:我们评估了在涉及不同认知过程和认知需求的3个决策任务以及另一个简单反应任务中,支配型动机对反应时的影响。
使用E-prime对行为实验进行编程。在选择任务时,通过在每个任务中涵盖不同的感官处理、认知负荷和决策过程,以非参方式探索反应时的潜在边界。并在任务间包含不同的指令和时间限制,以便如果发现与任务相关的反应时差异,我们可以探索有无时间限制是否起作用。
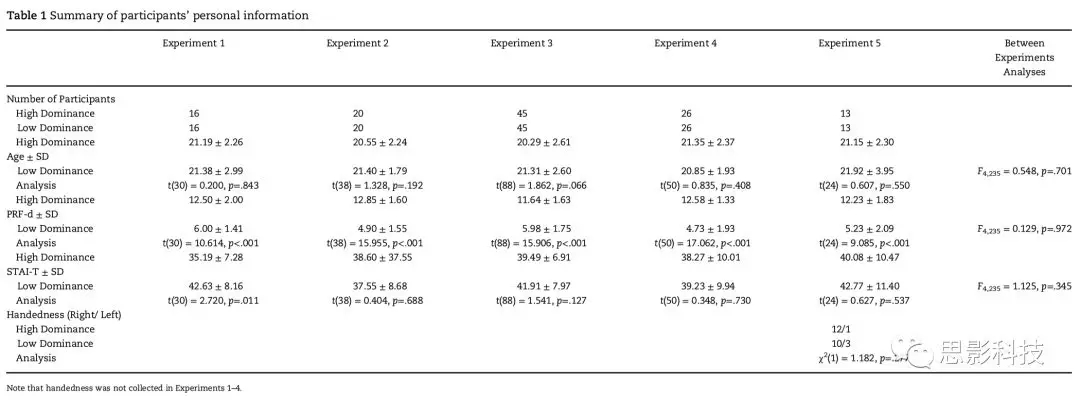
实验1是面部表情识别任务。实验刺激是男性脸部面孔的灰度图(72人),从thedatabases FACES, Nimstim Set of Facial Expressions, Radboud Faces Database和TheKarolinska Directed Emotional Faces数据库中获得。没有可以看见的衣服或珠宝并避免了引起注意的特征(例如疤痕、痣、面部毛发和不寻常的发型)。使用AdobePhotoshop CS5软件(版本12.0)将图像转换为灰度、均衡对比度、重新缩放图片到860×600像素,然后在保持原始纵横比的同时,手动标准化眼睛和下巴之间的距离。图像背景被设置为灰色,使用MatlabShine工具箱进行亮度均衡。向被试展示120张独特的图片:以其原始格式或翻转格式(通过镜像反转垂直轴上的原始图像获得)呈现每种情绪(愤怒、快乐或中性)的20种不同的面部表情。被试被要求注意中央“+”,并通过按下键盘键来选择呈现的是3种情绪中的哪一种。要求被试又快又准地做出反应。这些面孔在屏幕1/5的位置上呈现(以中心,20°或35°旋转到两侧),并且仅出现80ms以避免对目标进行扫视。在被试做出反应后立即呈现下一个面部图片(图1a)。这个实验评估了在决定面部表情时的反应时和正确率。
实验2是包括记忆和再认两个阶段的面部再认记忆任务。在第一阶段,要求被试记忆30张不同的面孔,其中15张和另外15张新面孔在第二阶段呈现,要求被试再认。我们使用了与实验1相同的图片,每张面孔呈现10s。为了探索与先前任务(被试在时间压力下执行)不同的时间限制,在这个任务中,被试可以在一定程度上以他们的速度进行。他们可以通过在记忆阶段按下空格键,或者在再认阶段回答是/否呈现下一个面孔图片(图1b)。
1)在记忆阶段之前,被试被告知在90s内尽可能多地记忆图片,并且在随后的再认阶段对他们的记忆进行测试。他们还被告知由于必须在90s内观看完30张图片,所以每张面孔平均允许呈现3s。实验记录到下一个图片呈现的时间。
2)在记忆阶段后5分钟内进行再认。向被试呈现面孔,并要求他们判断是否在第一阶段看到过该面孔。这一阶段没有反应时间限制,被试被告知在怀疑的情况下可以进行猜测。记录反应时和正确率。由于异常反应时剔除一个被试。
实验3是以工作记忆地图为基础的路线学习任务,旨在评估可能与组别相关的反应时差异。这次的任务与先前任务都不同,没有面孔和中等水平时间压力。此任务测验视觉工作记忆,先呈现地图轨迹,然后呈现测试轨迹,要求被试判断测试轨迹与地图轨迹是否相同。该部分结果是对Thoresenet al. 2016的数据进行重新分析,在重新分析中,根据PRF-D问卷对被试进行分层。在这一实验中,使用GoogleStatic Maps API(https://..google.com/maps)创建了48个具有相似路径密度的地图。地图面积为0.9km2,像素分辨率为640×640。其中一半地图是地图路线图,另一半是道路地图叠加的卫星地图。在设定路线之后被试观看红色标记的动画。每个试次包括一个学习刺激和一个测试刺激。在学习刺激中,标记进行17s,并告知被试学习路线轨迹。轨迹完成后,标记保持静态2s,并再次重复相同的轨迹。测试刺激开始,给被试呈现问题:“Isthis the same route as before?”,这一问题用法语呈现1s,并且在地图上以双倍速度呈现两次动画,尺寸减小27%。随机呈现试次,并且测试阶段的轨迹或者与以前一样,或者有细微差别的轨迹。被试在第二条轨迹出现时立即作出反应,但是他们被告知在观看重复的轨迹之后,他们将有4s的时间使用设定键来回答(“相同”或“不同”)。不要求快速反应,而且也不要求被试对所有试次作出反应。这项任务之前进行了四次预试验。分析第一个block中24个试次的数据的反应时和正确率(图1c)。
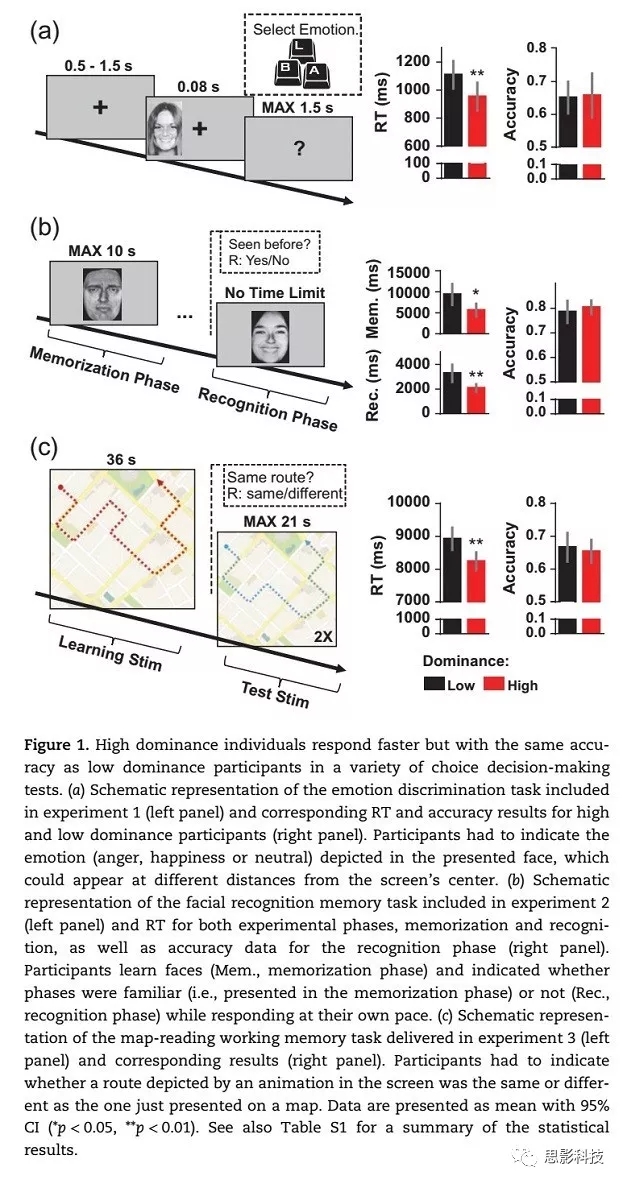
图1
实验4是简单反应时间任务。作为对照实验,该任务由交叉-非交叉差异任务改编(图2)。在每个试次中,灰色正方形出现在屏幕中央“+”的左边或右边。试次间的间隔在0.15到1.5s之间随机变化。要求被试一看到正方形,不管它出现在哪一侧马上按下空格键。0.5秒后,即使没有任何反应刺激也会消失。先进行六个试次的预实验,再依次呈现200个试次。每50个试次之间休息一次。
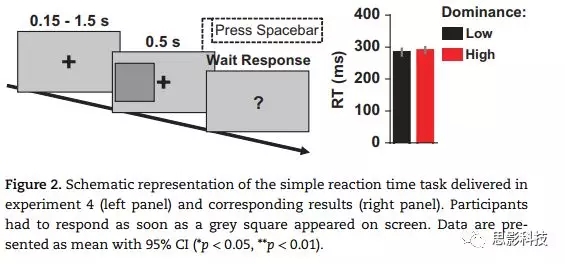
图2
EEG实验:
实验5旨在识别与高和低支配性被试反应迅速性差异相关的神经信号。为此,我们使用了实验1任务的改编版本。刺激包括40名男性或女性面孔(快乐,悲伤,愤怒,或中性情绪),并以随机方式呈现。从Ekman和Friesen的《Picturesof Facial Affect Series》中获得快乐和悲伤的面孔,而从FACES, RadboudFaces Database 和the Karolinska Directed EmotionalFaces中获得愤怒和中性的面孔。使用AdobePhotoshop CS5软件(版本12.0),将图像裁剪为面孔的中央部分,将亮度和对比度规则化,并将图片转换为相同大小(346×543像素)。所有的刺激都呈现在灰色背景上(RGB:192,192, 192)。在华硕VG248QE显示器上呈现图片,分辨率为1920×1080像素,刷新率为144Hz。在光线昏暗的法拉第笼中,被试坐在离显示器50cm处。背景亮度低于1cd/m2。注视通过眼动仪(The Eye Tribe)在整个实验过程中进行监测。头被下巴支撑杆固定。
所有被试完成两个实验条件,条件顺序伪随机分配给每个被试。在第一个条件中,刺激为快乐和悲伤的面孔(快乐与悲伤),而在第二个条件中,刺激为愤怒和中性的面孔(愤怒与中性)。每个条件有10个试次的练习,然后进行四个正式实验部分,每个部分80个试次。各实验部分之间间隔10s。要求被试又快又准作答,首先将视线保持在屏幕中央的“十”字上,并通过按键报告感知的情绪,当不确定时可进行猜测。在高支配和低支配的被试中,反应方和效价之间的联系是平衡的。每个试次中,在被试左侧或右侧26°呈现刺激0.1s,然后注视点呈现3s后被问号所取代,促使被试做出反应。在每个反应后,呈现0.5-1.5s的随机时间间隔。如果被试没有做出反应,将响起一个简短的嗡嗡声,在这一实验部分的末尾重复了这一试次。对于每个条件,正确率都是根据正确答案的百分比来计算的(图3a)。
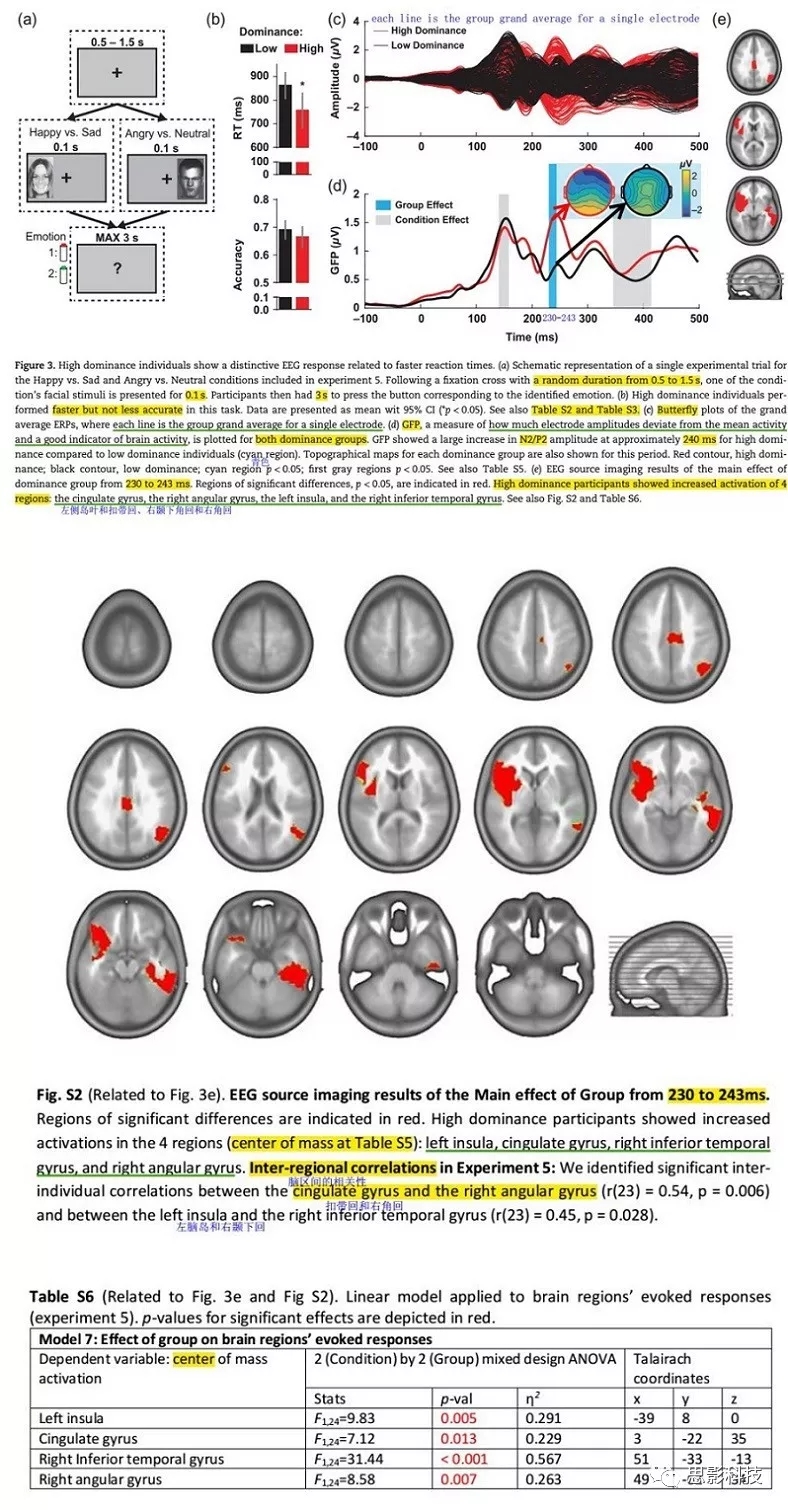
图3
EEG记录和预处理:
使用BioSemiActive 2系统(BioSemi)记录连续脑电图,该系统有192个Ag-AgCl烧结的活性电极,以共模感应(CMS)电极作为参考。调整帽子尺寸和位置:CZ电极位于枕骨隆突和鼻根点之间,电极均匀地覆盖整个头皮。记录眼电(EOG)的电极位于右眼上方和下方1cm处和外眦外侧1cm处。记录采样率为2048Hz。离线数据采样率降为512Hz,并使用自动预处理流水线(daCruz et al. 2018)处理离线数据。包括下列步骤:采用1-40Hz的带通滤波(3阶Butterworth滤波);去除线性噪声(CleanLine;www.nitrc.org/./cleanline);采用平均重参考;插值坏通道;去除坏段;采用独立成分分析(ICA)去除眼电、肌电和坏通道相关的伪迹;以及去除伪迹段。每个被试的插值电极的比例小于5%。我们从刺激前(基线)100ms到刺激后500ms选取时间段。每个被试的时间段都经过基线校正。每个被试的伪迹拒绝率都不到10%。
全局能量场分析(Global Field PowerAnalysis):
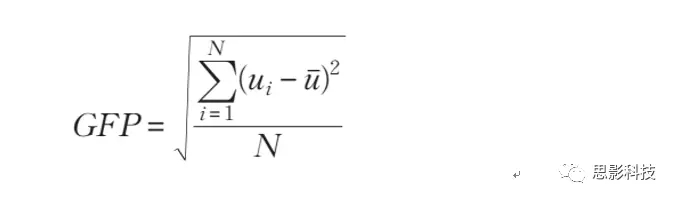
全局能量场(GFP)是一种瞬时的独立于参考的测量神经反应强度的方法,通过计算所有电极之间的电位标准差得到。考虑到时间自相关性,显著效应(p<0.05)连续持续10ms的才被认为是可靠的。分别计算每个被试、每个条件的GFP。每个时间点上的GFPs都采用重复测量ANOVAs,因素包括组别(低、高支配型)和条件(快乐vs悲伤,愤怒vs中性)。使用JeanFrançois Knebel开发的用于神经电成像的统计工具箱(STEN)来计算。
分布式源成像(Distributed ElectricalSource Imaging):
为了估计组别之间差异的大脑区域,我们使用局部自回归平均逆解(the Local Auto Regressive Average inverse solution)计算对应于GFP中显著组别效应的时间间隔逆解。使用CarTool软件进行源分析。使用STEN针对电流密度的每个解点,进行重复测量ANOVAs(组别[低、高支配型]×条件[快乐vs悲伤,愤怒vs中性])。使用以下空间标准对每个解点的多重比较进行部分校正:聚类必须包含至少15个显著效应的相邻解点(p<0.05)。为识别每个激活聚类中心(clustercenter of mass,CoM),对每个聚类显著区域上的电流密度进行平均。使用最接近CoM的解点活动来表示相应的大脑区域。
唾液皮质醇的分析(Salivary Cortisol Analyses):
收集3次唾液:1.在每个被试签署知情同意书之后;2.在洗漱之前,3.在第2个条件结束之后20min。在每次收集的10mL聚丙烯管中得到约0.8至1.4mL的唾液样品,并在-20℃以下冷冻直至加工。然后将样品在室温下以3000rpm的速度分离,持续15min,并根据制造商的指示(Salimetrics,Newmarket, Suffolk, United Kingdom)用酶免疫分析法测量唾液皮质醇浓度。样本被用来分析皮质醇基线水平和在实验过程中发生的激素变化。为了控制皮质醇的昼夜节律,所有实验安排在1pm和7pm之间。为了估计总体的皮质醇反应性,我们计算相对于基线的曲线下面积(areaunder the curve with respect to ground,AUCg)和相对于增加指数的面积(respectto increase indices,AUCi)。由于样本污染,5名被试被剔除,剩余21名被试(10名高支配型和11名低支配型)具有适合皮质醇AUCg和AUCi计算的完整的3组唾液样本。
数据分析:
行为数据:反应时小于200ms和超过规定时限的试次被剔除。反应时在所有被试平均反应时3个标准差以外的试次被剔除(每个被试所删试次不到5%)。如果存在被试内变量(在实验1中,情绪和难度为被试内变量),采用ANCOVAs或混合设计的ANCOVAs测量组别的反应时和正确率差异。当反应时是一个因变量时,正确率作为协变量以便解释这两个变量之间可能的权衡(除了实验4中包括一个与正确率无关的简单RT任务)。考虑到特质焦虑会影响个体的反应时,在分析因变量反应时和正确率时,ANCOVA将特质焦虑(用STAI-T评估)作为协变量。为了解释在EEG实验中被试间反应时的随机变化,使用了混合效应模型。固定效应被定义为“组别”ד条件”的交互作用模型,附加预测因子用来控制STAI-T和正确率。实验5的被试内变量为条件。随机效应是截距和斜率,它们分别解释反应时个体基线水平的差异以及由于条件导致的不同反应时变化。为了研究决策任务RT中PRF-d评分的整体效应,我们将实验1-5的混合效应模型与附加变量(编码决策实验)相结合。每个实验的RTs被标准化为Z分数,以避免比较不同时间尺度的反应。固定效应被定义为PRF-d×决策编码变量之间的交互作用,附加的预测因子用来控制STAI-T和正确率(如所有先前的模型所做的)。随机效应是随机截距(解释不同的实验)和随机斜率(解释实验2中的两个条件:记忆和识别)。lmerTest软件包采用自由度的Satterthwaite近似计算混合效应模型的p值。所有实验中报告的基于ANOVA方法的效应量为η2。用IBM-SPSS-StatisticsVersion 21.0和R编程环境进行统计分析。
皮质醇数据:皮质醇AUCg和AUCi指数作为ANOVAs的因变量,以评估在实验5中组别在糖皮质激素反应上的效应。
EEG数据:为了评估各脑区活动在RT或正确率上的效应,在因变量RT和正确率的独立回归中,显著的CoM激活被用作自变量。为解释RT在被试间的随机变化,使用了混合效应模型。固定效应是CoM激活,随机效应是与行为分析相似的截距和斜率。自由度的Satterthwaite近似被用来计算p值。pcor软件包采用皮尔逊偏相关计算显著CoM激活之间的被试间相关性。
结果及讨论:
高支配型个体比低支配型个体在决策任务中的反应更快:
在实验1-3(情绪识别任务[图1a]、面部识别记忆任务[图1b]、基于地图的路线学习任务[图1c];包括不同的认知需要和指令),我们评估了高、低支配型个体在反应时和正确率上的潜在差异。结果显示,高支配型被试比低支配型被试有更短的反应时,但两者正确率无显著差异。在实验4中(简单的反应时控制任务[图2])我们也评估了两组被试在反应时上的差异。结果显示,两组被试的反应时无显著差异。这一结论进一步得到单向贝叶斯ANOVA(one way Bayesian ANOVA)的支持,该分析描绘组别主效应的贝叶斯因子(BF10)为0.329,表明与其他假设(即,存在组间差异)相比,支持零假设(组间无差异)的证据增加了3倍左右。
我们的结果支持了假设,即使被试不在社会环境中,高支配型个体总是比低支配型个体更快。我们的策略是通过多个任务测试被试,以便能够覆盖广泛的情况,因此没有收集特定方面的参数操作数据。这种方法揭示了高支配型个体的快速反应贯穿于各种各样的任务中(实验1-3,实验5),因此,高支配型个体表现出更快的反应来应对不同程度的时间压力,包括情绪识别、面部学习和记忆以及地图阅读工作记忆。然而,高支配型被试在SRT中(实验4)没有优势的事实表明,他们的快速反应优势不是由于在感知过程或执行动作方面的优势。相反,高支配型个体表现出迅速反应的任务之间的共同因素是将选择和行动联系在一起的决策过程。
前人的发现表明,社会支配地位高的个体在竞争环境中特别敏感并激发他们的反应,特别是当面对竞争性挑战或发展出一般的支配感和优越感时(Balconiand Vanutelli 2016; Carver & White 1994; Gable et al. 2000; Hortensius etal. 2014;Hortensius et al.2014;Santamaría-García et al.2014; Santamaría-García et al. 2015)。然而我们的研究并未涉及社会竞争,但仍发现在选择决策任务中,高支配性个体的反应时仍短于低支配性被试。对于这种不一致的可能解释是,在我们的实验设置中所涉及的挑战对被试参与程度的影响随着被试的支配性动机的不同而不同。或者由BAS问卷(前人研究中使用)和PRF-d问卷(在我们的研究中使用)定义的高支配性被试可能不完全重叠。使用PRF-d量表,我们可能获得的被试表现出一种特别投入的认知风格,只关心决策情境中行动的迅速性,而不管是否具有偶然的竞争性质。未来的研究应该包括通过不同的研究来交叉验证发现的支配性测量。
有趣的是,我们的行为发现与动物方面的文献一致,这些文献表明主动(比他人的反应更勇敢,更具攻击性)行为类型与社会支配性和竞争能力之间存在着正相关(Davidet al. 2011; Harcourt et al. 2009; Mamuneas et al. 2015; Nakayama et al. 2012; Oortmerssenet al. 1984; Riebli et al. 2011)。在人类中,支配性也是领导能力的一个强有力的预测因子。因此,我们的研究结果可能对领导力的产生有一定的影响。根据在许多动物社会结构中运作的领导者-追随者决策规则“跟随先行者”(leader-followerdecision rule “followthe one who moves first”),人们认为成为领导者的可能性取决于特定的内部或社会特征,这些特征增加了行动开始的可能性。这一现象暗示着反应时是社会支配性的指示性心理测量变量。在涉及资源控制或确保生存的决策情况下迅速行动可以提供进化优势并促进支配行为的出现。在没有速度-准确性权衡的情况下,支配性个体更快的认知过程可能有助于指导群体决策,而不会损害准确性。最近,反应的迅速性被证明与同伴对个人魅力的感知增加有关。
高支配型个体表现出独特的EEG信号:
为了识别与高支配型个体的快速反应相关的脑激活,我们用高密度EEG进行了实验5,被试完成修改后的实验1中的情绪识别任务(图3a)。结果发现,高支配型被试的反应时比低支配型被试短(图3b;β= 127.57 ms, SE = 60.88 ms, 95% CI[5.82, 250.89], t(23.59)=2.10, p = 0.047)。正确率无显著差异(图3b)。我们还证明了,当PRF-d分数被看作连续变量而不是上面所使用的二分法时,在所有决策实验中都发现了与支配型相关的反应时效应。该结果与实验1-3的结果一致。
鉴于以前的研究表明糖皮质激素水平的提高可以影响神经回路的激活,我们在实验的不同时间点测量唾液皮质醇。在整个实验中,我们发现各组间皮质醇水平无显著差异(AUCg:F1,19 = 0.23, p = 0.638, η2 = 0.012, 95% CI[<0.001, 0.070]; AUCi: F1,19= 0.24, p = 0.633, η2 =0.012, 95% CI [<0.001, 0.072])。
尽管一些研究强调了皮质醇与竞争环境中的社会支配地位之间的联系(Mehtaet al. 2008; Mehta and Josephs 2010; Turan et al. 2015),但是在我们的研究中,高和低支配组的皮质醇水平没有显著差异。考虑到我们的实验不涉及竞争,并且每个被试都单独测试他们的行为反应,皮质醇水平缺乏社会支配性相关的差异与文献完全一致,表明皮质醇差异仅出现在社会竞争挑战中(Mazuret al. 2015; Larrieu et al. 2017; Turan et al. 2015; Wirth et al. 2006)。因此,我们的发现排除了在皮质醇作用上,与社会支配性相关的大脑活动差异。然而,我们不能排除影响大脑活动动态性的其他压力相关系统(例如,大脑去甲肾上腺素),他们在这两种支配性群体的任务表现中可能会有不同的参与。
刺激呈现后大约210到280ms处(前部N2和后部P2成分),高支配型被试诱发了更显著的偏转(参见图3c中的蝴蝶图)。GFP的重复测量ANOVA显示,两组在刺激呈现后230-243ms之间有显著差异(F1,24= 4.87, p = 0.037, η2 =0.169, 95% CI[0.006, 0.370],图3d)。高支配型被试比低支配型被试有更大的偏转。各实验block之间有显著差异,但“组别”ד条件”的交互作用不显著。
EEG中的N2/P2成分可以反映对目标刺激觉察、分类和决策相关过程的资源分配,以及执行任务的意志努力。因此,在N2/P2成分上,与支配性相关的差异可能反映了调动这些资源方面的缺陷。作为推测,N2/P2响应的显著增加可能是社会支配性的一般指标。在大约240ms,识别大脑过程的内在差异可以定义为认知风格的关键差异,当在社会环境中时,这些差异为支配性等级和领导――追随者关系的出现奠定了基础。
在定义的组别效应窗口(230-243ms),EEG源成像分析显示相对于低支配型个体,高支配型个体在扣带回、右角回、左脑岛和右颞下回的活动增加(图3e)。在探索性分析中,我们测量了扣带回活动与反应时之间的关系,以及扣带回与右角回、左脑岛与右颞下回之间的个体间相关性。
以前的研究将扣带回的高度激活与更快的反应时联系,尤其是当刺激位置不可预测时。如在我们的实验中,面孔被呈现在注视点的左侧或右侧。扣带回激活也与运动准备和注意力分配方面的认知努力增加有关。因此,扣带回皮层的较高激活可能反映高支配性个体有更高的能力招募大脑资源以促进反应选择。此外,高支配性被试表现出在执行任务中激活区域的增加。具体而言,右侧颞下回包含参与面部过程的梭状回面孔区域,并且在情绪感知和体验过程中观察到了左脑岛的活动。
未来研究方向:
在EEG研究中,早、中或晚期成分(如P1、N1、P3或单侧化准备电位)可能出现群间差异。早期的差异可能被视为更快的感官编码和视觉处理的证据。P3的后期差异可能已被视为认知差异的证据,与支配性以及更快动作执行的准备电位的差异有关。如前所述,我们仅发现中期成分的显著差异,表明高支配性个体将感官证据更快速地转化为决策。我们的发现产生了几个问题:1)确定N2/P2响应是否容易改变,例如,当一个高支配性个体具有与上述结果类似的下属角色时。2)社会等级已经在学龄前儿童中被观察到了,因此,需要评估在发展阶段的哪一时期出现增加的N2/P2成分。3)把我们的发现运用到现实生活中。4)女性被试是否会表现出类似的行为和EEG模式。5)确定我们能否发现快速反应时、识别出的大脑区域活动增加、以及日常生活参数(如收入、社会地位、体育运动)之间的相关性。
作者提出研究结果本质上是相关。因此,不能对因果关系作出任何结论。例如,虽然快速反应可能有利于生存,但是它们也可能只是支配性的表现,类似于支配性个体在群体中率先发表意见。同样地,我们不能确切地知道大脑活动的增加是否是导致快速反应的原因,或者仅仅反映了与支配性有关的其他特征类型的加工过程。此外,我们不能排除这种可能性,即与支配动机有关的权力感和优越感会影响任务的参与度,因此,本研究的ERPs和反应时类似于最近发现的人工等级分配对反应时和ERPs的影响。
在实验室任务中发现的快速反应的加工速度,被认为是一个强有力的生存预测因子,而且更高等级的个体表现出更健康的状态。令人惊讶的是,尽管健康和幸福的很多优势与高社会等级相关,但对于那些使个体倾向获得支配地位的因素,我们知之甚少。我们的研究提出了一种可能性即在决策情况下的快速反应差异,以及相关神经基础的差异是等级建立的核心,并且可能将社会等级与身心健康联系起来。
da Cruz, J., Rodrigues, J., Thoresen, J. C.,Chicherov, V., Figueiredo, P., Herzog, M. H., & Sandi, C. (2018). Dominant men are faster in decision-making situations andexhibit a distinct neural signal for promptness. Cerebral Cortex.

欢迎微信扫码或者长按二维码关注我们
思影科技给您带来最新脑影像资讯
获取原文:关注“思影科技”公众号,回复“原文”或“培训”,获取原文pdf及补充材料下载链接,同时欢迎浏览我们的培训通知以及数据处理业务介绍。(直接点击下文橙色字即可浏览):
第九届脑电数据处理基础班
第三届脑电信号数据处理提高班
第一届眼动数据处理班
第十届功能磁共振数据处理基础班
第四届近红外脑功能数据处理班
第六届磁共振脑网络数据处理班
第二届动物磁共振脑影像数据处理班
第五届磁共振弥散张量成像数据处理班
第四届磁共振脑影像结构班
第三届磁共振ASL(动脉自旋标记)数据处理班
思影数据处理业务一:功能磁共振(fMRI)数据处理
思影数据处理业务二:结构磁共振成像(sMRI)与弥散张量成像(DTI)处理业务
思影数据处理业务三:ASL数据处理
思影数据处理业务四:EEG/ERP数据处理