大脑的出色语言能力要求特定功能脑区之间相互影响。荷兰Radboud大学的Schoffelen等人在PNAS发文,P这一结果与这些脑区在语言处理中扮演中心角色的观点非常吻合。
采用数据驱动将矩阵分解出一系列子网络;由该方法获得的功能脑网络拓扑属性揭示了不同频率相互作用下的有向连接。来自颞部的连接在 α 频率时达到峰值,而来自额叶和顶叶的连接在 β频率时达到峰值。这些发现表明,语言处理所必需的与语言相关的脑区之间的信息流动可能取决于不同的大脑节律所起的作用。
研究背景:
脑电图和脑磁图(EEG/MEG)的电生理研究表明,语言网络中各个节点的序列激活具有较高的时间精度。但是使该网络各节点之间的信息能够有效流动的功能交互的性质尚未阐明。
皮层脑区间连接的特征是信息可以双向交换。同步脑区节律可能促进大脑皮层区域间的信息流动,并且特定的不同频率的神经元节律反应了信息流动的方向。这种双向的信息流动是支持语言处理的神经生物学系统的一个重要特征。语言处理(eg:阅读句子)不是一个简单的bottom-up的过程;由上下文信息也可以提供top-down的信息。
在这里,作者发现与语言相关的脑网络中,由特定频率下有节律性的神经元同步可以促进脑区间的相互作用。具体地说,作者发现 α ( 8-12 Hz )的节律性活动是由颞叶皮层传播到额叶皮层,而 β ( 15-30 Hz)的传播方向相反。这些结果表明在语言处理过程中,有节律的定向交互的功能相关性取决于由不同频率段支持的脑区间相互作用的方向。这种功能相关性可扩展到其他认知领域,即反映一种与任务相关的脑网络中的信息动态流向的通用机制。
方法
1、实验过程和MEG数据采集
被试以荷兰语为母语( n=102, 其中51名男性),年龄在18-33岁之间。所有被试都是右利手,视力正常或矫正后正常,没有神经、发育或语言缺陷史。将要阅读的句子和单词序列(共240个单词,每个单词序列9-15个单词)依次呈现在被试前的反投影屏幕上。MEG数据采集频率为1200 Hz。在整个测量过程中,头动信息使用软件监视。
2、去伪迹
所有分析都是通过 MATLAB 脚本和FieldTrip完成。由单词开始计时,数据由 -100 ms到600 ms 进行分段;并且在进一步分析前去掉干扰伪迹的分段。使用ASEO 算法从单试次数据中去掉事件相关响应,以减弱诱发瞬态对估算 GC(格兰杰因果关系)的影响。由于各脑区瞬态信号峰值的系统延迟差异,使得这些信号的转变违反了静态假设,并导致GC中出现非零值。使用 ASEO算法和盲源分离技术结合,将单次试次信号建模为正在进行的活动和事件相关的成分的组合,并从数据中去掉在模型中单次试次引起的响应。
3、 源重建和重建源活动的分割
使用 LCMV 对源进行重建,即对 8196 个位置估算空间滤波。然后,采用了一种基于atlas的分割方案来降低数据维度。使用Conte69 模板将每个大脑半球分割成191个脑区。
4、 预先选择语言相关脑区之间的连接
根据前人文献的描述,我们使用核心语言脑区和视觉系统构成的脑区之间的已知“长程”宏观解剖纤维通路,先验地选择研究一些脑区之间的连接。根据皮质-皮质连接的特点,我们会先验的进行邻近节点之间的有向连接。既包括了来自两个大脑半球的半球内连接,也包括同源区域之间的半球间连接。
Fig.2A显示了各个节点的标记方式。在成对的脑区间产生了4350个连接,其中包括用于GC(格兰杰因果关系)分析的156个脑区间所有可能成对连接。
5、 整个网络拓扑结构的GC计算和统计
计算出频谱信号,并使用特定的空间滤波器将其投射到源空间。利用快速傅里叶变换和multitapers(5Hz平滑)法结合,将单词呈现后的200-600ms时间段的时域信号转换为频谱信号。计算每一对脑区间的交叉谱密度矩阵,并利用它来计算GC。此外,还根据时间反转数据的源投射傅里叶变换计算GC。只选择GC和反向GC的差异具有统计学意义的脑区对进行统计分析,Bonferroni多重比较校正(p<0.05)。这将用于后续分析的连接数量从4350个减少到713个。
6、NMF算法和网络可视化
通过对生成的 GC (格兰杰因果关系)频谱进行稀疏约束的 NMF(稀疏非负矩阵因子分解) 来研究网络拓扑结构。本分析的目的是将重建的连通性数据描述为网络成分的低维混合,其中每个成分都具有特定被试的频谱剖面。选择稀疏非负矩阵因子分解(NMF)是因为非负性约束有助于解释成分。
NMF的结果依赖于成分的数量。本研究确定了 20 个成分数,在提供少量可解释成分的同时,保持子网络之间的较好分离,从而达到合理的平衡。忽略 NMF 算法的随机初始化效果,使用重复随机初始化40次的LCASSO框架对结果聚类的过程来提取数据的底层结构。
7、特定条件下的统计分析
由已识别的网络成分提取出的主要连接,估算特定条件下的GC。条件是根据单词是否出现在句式结构良好的句子上下文(或单词序列的一部分);以及单词是否出现在句子(或单词序列)前段(2-4个单词)或句子(或单词序列)后段(n-3到n-1,n是语句序列的总词汇数)来定义。在本研究中,保证了在不同条件下时域信号差异和词汇频率边缘分布的平衡性。由于脑区对特定的分段数量因脑区对有差异,因此只用一个分段子集来计算GC。最终保留了147个分段。计算特定被试和特定条件的GC,并进行了非参数检验来评估:
(i) 句子单词 VS 单词序列单词;
(ii) 句子条件:前段单词 VS后段单词,
(iii) 交互效应:
(前-后单词)句子-(前-后单词)单词序列。
结果
使用 MEG 记录被试阅读句子时的脑磁信号。在一组预定义的脑区(包含156个皮层区域)中重建皮层活动。这些脑区包含了核心语言系统、视觉系统在内,以及大脑对侧半球相同位置的区域(如图 1A,即 Fig.1A )。
接下来,计算GC 来量化与语言相关的脑区之间的定向节律性神相互作用。由脑磁图记录估算得到的连通性的解释在空间上的分辨率很低。作者对102名被试信号的时间反转前后的GC估算值进行了统计比较。保守丢弃了大量预定义的连接子集,最终保留来自预定义的156个皮层区域间(4350个连接)的713个连接子集。随后,研究了得到的网络的拓扑结构,并观察到所涉及的皮层区域的连接数分布不均匀(图1 B和C;即Fig.1B&C)。具体地说,量化了每个皮层区域的向内和向外连接的数量(如,节点度)。
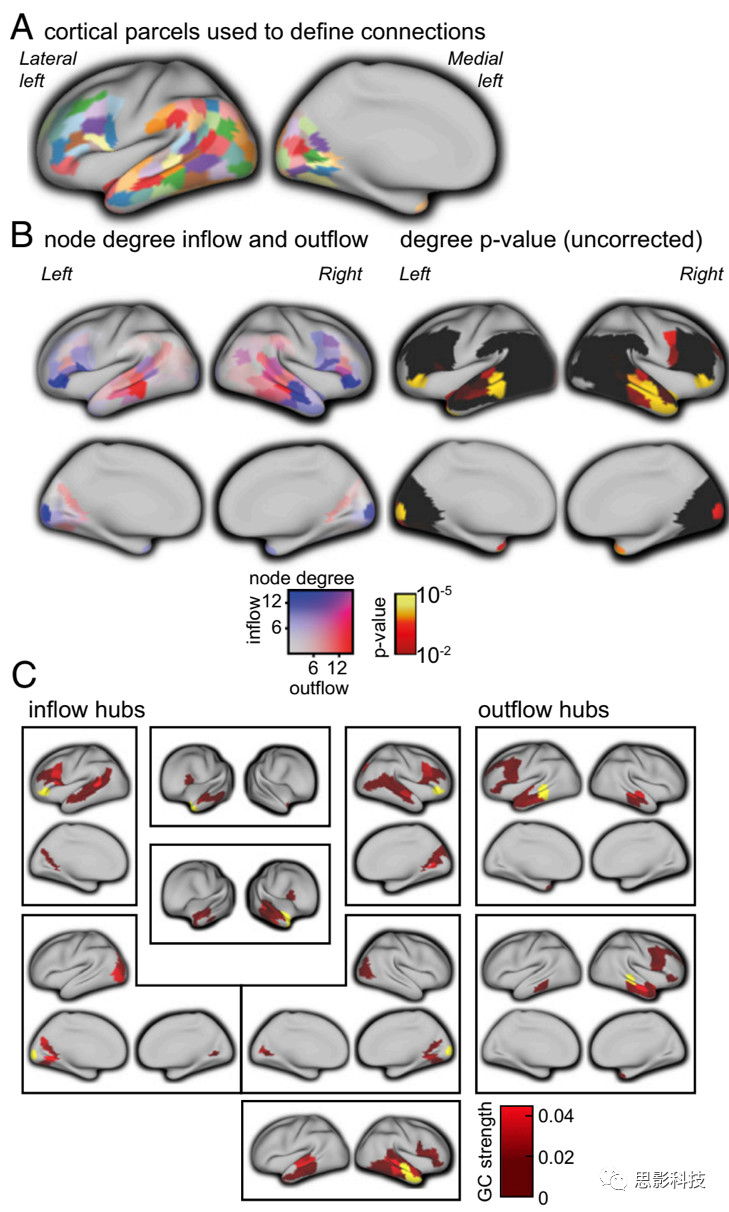
图1 用GC对语言脑网络的拓扑结构进行量化研究。
(A)左半球解剖脑区,用于源重建和作为网络节点,展示在膨胀的皮层表面。左图和右图分别显示外侧和内侧表面。右半球对称的脑区也被考虑用于网络估计,但这里没有显示。
(B左图)信息流入和流出的节点度。
(B右图)未校正的p值,以及橙色/黄色经过Bonferroni矫正存活的脑区。
(C)B中发现的脑区和其他皮层脑区的拓扑连接结构,换色是B中脑区的信息流入(蓝色)和流出(红色)脑区,GC强度用红色表示。
作者采用Bonferroni多重比较矫正(p<0.05),得到左右颞中皮层区域作为大量连接的发送节点;并由它投射到同侧的前中颞上皮层(Brodmann 分区 (BA) 21/22/38)、对侧的中颞上皮层(BA 21/22),以及额叶皮层(BA 6/9/44/45/47)。同样采用Bonferroni矫正(p<0.05),得到左右额叶脑区(BA 47)接收到来自同侧额叶皮层(BA 44/45/46)、同侧颞上皮层(BA 22)、同侧角回(BA 39)和同侧纹外视觉皮层( BA 19,17/18号脑区只出现在大脑右半球)的格兰杰因果输入。此外,还发现接收信息大量流入的脑区位于双侧前颞极、枕极和右前颞叶皮层。
为了更详细地了解这个脑网络的空间和频谱结构,作者将NMF(稀疏非负矩阵因子分解)应用到组水平连接数据。具体操作是,将连接数据建模为有限的空间静态网络类别的混合;其中每个类别都具有被试特定的频谱剖面。其分解算法没有对边缘进行空间上的聚类(即脑区间的定向连接在空间上聚类时,分解算法不支持连接集聚在同一类别中)。但是,聚类得到的网络类别大部分在生理上具有可解释性。
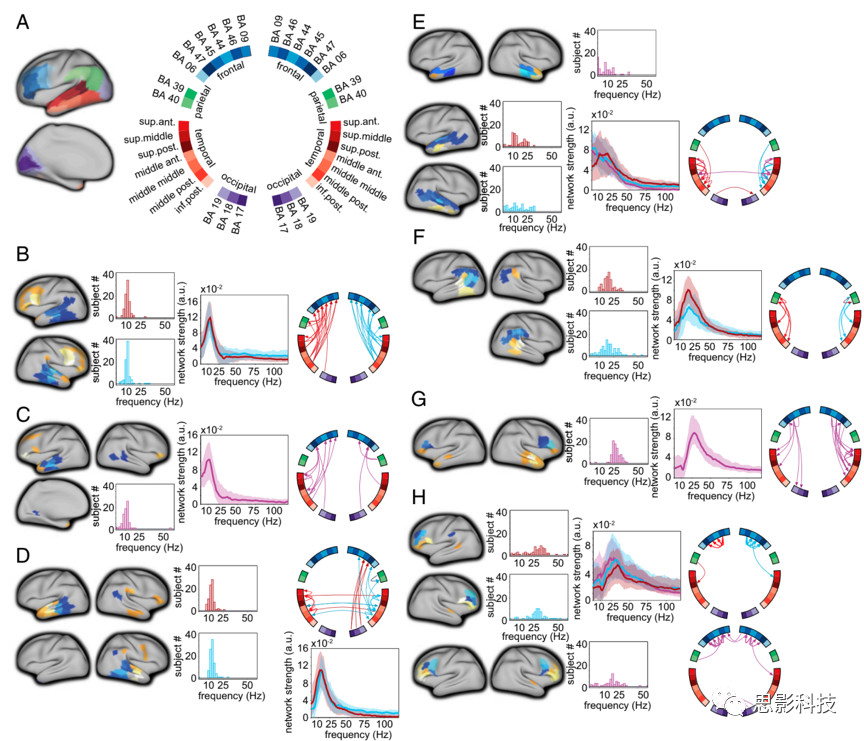
图2 NMF计算出的网络成分,图中显示了频率,脑区,定向的交互。
(A)脑区节点的定位展示在膨胀后的皮层上,有颜色编码(左图)和标签(右图)。Circular图用的是布鲁德曼标签。
(B-H)语言相关脑区的脑网络成分图,深/浅蓝色:信息流出的脑区(颜色越深,这个脑区对网络成分的贡献越大)。橙色/黄色/白色:信息流入的脑区。柱状图显示了每个网络成分的特定被试的峰值频率分布。频谱图用的是中位数(不是均值)对频谱情况的描述。Circular图显示的是脑区间的有向连接。箭头的厚度反映了连接的相对强度(反正我看不出区别)。以左半球、右半球或双侧连接为主的网络成分分别显示为红色、淡蓝色和紫色。
(B)左右半球成分从颞区到同侧额叶区。
(C)双侧半球内颞额连接。
(D)左右半球成分中的主要连接是从中部到前颞区。
(E)由上至下脑区(上两个)和由中前区域到颞极(底下那一行)的颞叶内的主要连接。
(F)角回(BA
39)和缘上上回(BA 40)与后颞叶的主要连接。
(G)额叶到颞叶的主要连接。
(H)主要是额叶的连接。
Fig.2是与语言相关皮层脑区间主要连接的网络类别。对于某些类别,被试的平均频谱被限制在一定的频率范围内;而且在被试间呈现出一致的峰值频率(图2 B-H,即 Fig.2 B-H;对应于四列的中间位置示意图)。此外,还确定了左右半球从后颞叶皮层和中颞叶皮层到同侧额叶皮层(主要是下额叶)的定向节律相互作用;其中峰频率为 12
Hz(四分位距(IQR)11-13 Hz)(见Fig.2B)。如 Fig.2 C 所描述的是一个在空间上主要由左半球内连接的较为弥散的从颞中区通向下额叶和额叶上区的类别。如 Fig.2 D所示,与颞叶-额叶间的连接相比,颞叶后区和颞叶中区到同侧颞叶前区皮层的连接具有略高的中位数峰值频率,14Hz(左右半球类别的 IQR分别是12-15 Hz和13-15 Hz)。 Fig.2
E 所示的是,一组主要相互连接的颞叶皮层脑区的频谱在被试中表现出更大的变异性。这些类别反映了由上、中颞皮质(沿整个前后轴)到中、前下颞叶皮层的连接;以及中、上颞叶前部皮层到颞极的连接。
与颞叶皮层中输出脑区的网络类别相比,由顶叶(Fig.2F)和额叶(Fig.2G,Fig.2H)为主要流出的脑区的节律性交互始终表现出较高的交互峰值频率。聚类的类别反应出由颞顶叶到后颞叶的交互平均峰值频率为20 Hz(左右半球类别的IQR分别是17-22
Hz和15-26 Hz),额叶到颞叶的节律性交互平均峰值频率为27
Hz(IQR是25-30 Hz)。在BA的44号脑区到BA的45/46/47号脑区的定向交互中,中峰频率为24 Hz,峰内的相互作用具有较宽的频谱范围。由BA 46号脑区到44/45/47号脑区的交互作用中,左右半球聚类类别的中峰频率分别为30 Hz (IQR为23-35 Hz)和 29 Hz (IQR为25-33 Hz)。
Fig.3A 描述了顶叶、额叶和颞叶间的主要连接的聚类类别之间节律性交互的峰值频率。总体而言,特定类别的中峰频率范围由 α 频率段上限 (12
Hz) 到 β 频率段上限值 (30 Hz)。此外,通过置换检验(p<0.05)得到,以颞叶为主的节律性格兰杰因果流出的聚类类别的峰值频率始终比顶叶或额叶流出的更低。值得注意的是,基于NMF(稀疏非负矩阵因子分解)可以区分颞叶到额叶的交互和由额叶到颞叶的交互。前者的峰值频率为12 Hz,用深红色标示(Fig.2A, 2B和Fig.3B, 3C);后者的峰值频率为27 Hz,用深蓝色表示(Fig.2F和 Fig.3B,3C)。其中 Fig.3B 为主要节律相互作用的示意图,Fig.3C为相应的频谱图。

图3 颞顶叶皮层的节律性相互作用的峰值频率始终低于额叶皮层的峰值频率。
(A)网络成分特定的峰值频率的两两比较(非参置换)。每个颜色表示被试特定的峰值频率差异的中位数。沿着主对角线的黑方块中的值反映了每个成分的峰值频率的中位数。
(B)语言系统中定向的节律性的脑区相互作用的示意图。颞叶分为两个节点,能够显示节律性的中到前脑区的连接。彩色箭头指的是C图中的频谱。
(C)C中的黑线是图2E中各成分的平均的功率谱。主要是颞上区到颞中回的连接。
接下来的研究是,节律性交互的强度是否受到感知输入所施加的功能需要的调节。将刺激材料分为 4种情况:
被试阅读的结构完整句子,由单词构成的伪随机序列(单词序列),单词的顺序位置是出现在前段,单词顺序出现在序列后段。
通过在空间上聚类的方法,从NMF结果中提取每个被试的格兰杰因果交互作用和最显著的功能连接的条件。并对频率和边缘进行平均。由各聚类类别的峰值频率和IQRs获得特定连接的频率段。将完整句子和单词序列比较,经过非参置换检验(P<0.05)Holm-Bonferroni多重比较校正发现,从左中颞叶到左颞极的相互作用强度被调制;从右纹状到纹外视觉皮层的作用强度单词序列要大于句子的,见Fig.4A。Fig.4B是早期(单词在序列前段出现)、晚期(单词在序列后段出现)单词间的比较结果。结果显示,在句子中,早期有更强的节律性相互作用。从颞叶到额叶、颞叶中区到颞极的连接是双侧的。此外,在右半球中发现,额叶到颞叶、颞上回到颞中回之间存在显著的调制连接(置换检验,p<0.05的多重比较校正)。

图4 有向交互的强度受语言输入的调节。
(A)在单词列表条件下,连接显示出比在句子条件下更强的交互(P<0.05)。蓝色脑区是信息流出,黄色是流入。
(B)与句子中较晚出现的词相比,句子中较早出现的词之间的联系显示出更强的相互作用。红色框里显示较强的交互效应。
(C)柱状图显示了不同条件的GC强度的均值。
讨论
本研究为与语言相关的脑网络皮层区域在阅读句子时的定向交互提供了证据。对整个网络的拓扑分析显示,大量的格兰杰因果流入额前下皮层区域、右侧颞前皮层和双侧颞极。
颞中皮层区域表现出高度的格兰杰因果流出。值得注意的是,没有在连接模式中观察到明显的双侧化,这说明大脑的两个半球都参与了语言刺激的处理。
数据驱动将整体网络分解为更小的子网络,揭示了几个对应于局部和长程定向交互的在空间上受限的类别。来自颞叶皮层区域的连接峰值频率( α和低的β 频率) 始终低于来自顶叶或额叶区域(高β值)的连接峰值频率。此外,本研究的数据还揭示了大脑语言系统中特定频率下的子网络。
进一步探索这些交互作用的潜在功能意义,发现语言环境调节左外侧颞中-前颞间的相互作用,以及右外侧纹外皮层-纹状体相互作用。在这些连接中,格兰杰因果影响在由单词构成的序列条件下更强。这表明了句子结构的缺失加剧了信息传递到前颞叶的需要。此外,研究结果表明,在没有上下文约束的语言环境下,即使是单词形式,知觉的产生也可能需要更强的交互作用才能有效进行处理。
综上所述,本研究显示了语言相关脑区的高度动态皮质网络中的定向交互作用,在支持颞叶-额叶和额叶-颞叶方向信息流动的特定频率上存在显著差异。虽然研究结果与之前关于正反馈和负反馈连接之间频率差异的报告一致,但这里显示的语言网络中的频率与在视觉系统中观察到的频率有所偏离。语言环境对其中一些连接强度的影响表明了一般认知过程中动态节律性皮质相互作用的功能相关性,尤其是语言处理过程。进一步研究的一个途径是将这些节律性的相互作用与局部的激活方式联系起来,从而深入了解局部皮层活动和长期相互作用之间的相互作用是如何塑造认知的。
如果您对格兰杰因果分析和脑网络分析以及EEG高阶功能连接感兴趣,请点击以下链接浏览:
思影数据处理服务六:脑磁图(MEG)数据处理
第五届任务态fMRI专题班(南京)
第十二届磁共振脑网络数据处理班(南京)
第五届脑电信号数据处理提高班(南京)
总结
大脑的出色语言能力需要特定功能脑区之间的双向作用。尽管目前已很好地研究了大脑中各个脑区在语言网络中的功能作用,但是对于促进这些脑区间信息交换的机制却知之甚少。在这篇论文中,证明了大脑中与语言相关的脑区之间的作用是由有节律性的神经元同步促成的,不同的节律反映信息流的方向。这些发现可能反映了一种在认知处理过程中,允许与任务相关脑区中的信息动态流向的通用机制。
原文:
Frequency-specific
directed interactions in the human brain network for language
JM Schoffelen, A
Hultén, N
Lam
如需原文及补充材料请加微信:siyingyxf 获取,如对思影课程感兴趣也可加此微信号咨询。

微信扫码或者长按选择识别关注思影
感谢转发支持与推荐
欢迎浏览思影的其他课程以及数据处理业务介绍。(请直接点击下文文字即可浏览,欢迎报名与咨询):
第五届任务态fMRI专题班(南京)
第十二届磁共振脑网络数据处理班(南京)
第二十三届功能磁共振数据处理基础班(南京)
第十届磁共振弥散张量成像数据处理班
第一届弥散磁共振成像数据处理提高班
第八届磁共振脑影像结构班(南京)
第八届脑影像机器学习班(南京)
第二十五届磁共振脑影像基础班(南京)
第七届脑影像机器学习班(重庆)
第二十六届磁共振脑影像基础班(重庆)
第六届磁共振ASL(动脉自旋标记)数据处理班
第五届小动物磁共振脑影像数据处理班(重庆)
第二十四届磁共振脑影像基础班(重庆)
第五届脑电数据处理入门班(南京)
第十八届脑电数据处理中级班(南京)
第五届脑电信号数据处理提高班(南京)
第十七届脑电数据处理班(重庆)
第五届眼动数据处理班(重庆)
第六届近红外脑功能数据处理班(上海)
思影数据处理业务一:功能磁共振(fMRI)
思影数据处理业务二:结构磁共振成像(sMRI)与DTI
思影数据处理业务三:ASL数据处理
思影数据处理业务四:EEG/ERP数据处理
思影数据处理服务五:近红外脑功能数据处理
思影数据处理服务六:脑磁图(MEG)数据处理
招聘:脑影像数据处理工程师(重庆&南京)