―Γ‘ώ–‘ΉΔ“βΩ…“‘Ά®ΙΐΉΔ“β‘ω“φ”κΫΒΒΆ…ώΨ≠‘κ…υά¥‘ω«ΩΕ‘Η–Ψθ–≈œΔΒΡ¥ΠάμΓΘ»ΜΕχΘ§’βΝΫ÷÷Μζ÷Τ‘ΎΕύ¥σ≥ΧΕ»…œ”–÷ζ”ΎΗΡ…ΤΉΔ“βΙΐ≥Χ÷–ΒΡ÷ΣΨθ±μœ÷»‘»Μ¥φ‘Ύ’υ“ιΓΘ±ΨΈΡΦΌ…ηΘΚ≤…”ΟΡΡ÷÷―Γ‘ώ–‘ΉΔ“βΜζ÷Τ»ΓΨω”Ύ»ΈΈώ―ΒΝΖΒΡ≥÷–χ ±ΦδΓΘ±Ψ―–ΨΩΆ®Ιΐ“ΜœνΒδ–ΆΒΡ―Γ‘ώ–‘Ω’ΦδΉΔ“βERP Β―ιΖΕ ΫΘ§Ψ≠Ιΐ1Ηω‘¬ΡΎ20¥ΈΡ‘Βγ≤β ‘Θ§≤…”ΟœΒΆ≥ΕχΖαΗΜΒΡ¬έ÷ΛΘ§ΒΟΒΫ÷Ί“ΣΒΡΫα¬έΘΚΉΔ“β‘ω“φ‘Ύ―ΒΝΖ‘γΤΎΤπ÷ςΒΦΉς”ΟΘ§ΒΪ‘Ύ―ΒΝΖΚσΤΎΤπ÷ςΒΦΉς”ΟΒΡ «…ώΨ≠‘κ…υΫΒΒΆΓΘ’β“ΜΙέ≤λΫαΙϊΕ‘”ΎάμΫβΉΔ“βΜζ÷Τ“‘ΦΑΆΤΙψ Ι”Ο≤ΜΆ§ΡΘ–ΆœΒΆ≥(άΐ»γΘ§»ΥάύΚΆΖ«»ΥάύΝι≥ΛάύΕ·Έο)ΒΡ―–ΨΩΫαΙϊΨΏ”–÷Ί“Σ“β“εΓΘ±Ψ―–ΨΩΖΔ±μ‘Ύ÷χΟϊ‘”÷ΨΓΕPLOS Biology ΓΖ…œΓΘ
ΈΡœΉΒΦΕΝ
―Γ‘ώ–‘ΉΔ“β÷ß≥÷œύΙΊΗ–Ψθ–≈œΔΒΡ”≈œ»¥ΠάμΘ§“‘¥ΌΫχΡΩ±ξΒΦœρ––ΈΣΓΘΕ‘»Υάύ≤Έ”κ’ΏΒΡ―–ΨΩ±μΟςΘ§¥σΡ‘ΤΛ÷ Ζ¥”ΠΒΡΉΔ“β‘ω“φΩ…“‘≥δΖ÷ΥΒΟς”κΉΔ“βΝΠœύΙΊΒΡ––ΈΣΗΡ…ΤΓΘΝμ“ΜΖΫΟφΘ§ Ι”Ο―ΒΝΖ”–ΥΊΒΡΖ«»ΥάύΝι≥ΛάύΕ·ΈοΫχ––ΒΡ―–ΨΩ±μΟςΘ§ΫΒΒΆ…ώΨ≠‘κ“τΩ…“‘ΗϋΚΟΒΊΫβ ΆΉΔ“β¥ΌΫχ––ΈΣΒΡ‘≠“ρΓΘΦχ”Ύ―Γ‘ώ–‘–≈œΔ¥Πάμ‘ΎΦΗΚθΥυ”–»œ÷ΣΝλ”ρΒΡ÷Ί“Σ–‘Θ§Έ“Ο« ‘ΆΦΆ®ΙΐΓΑ≥ΛΤΎ––ΈΣ―ΒΝΖΜαΗΡ±δ÷ß≥÷―Γ‘ώ–‘ΉΔ“βΒΡ…ώΨ≠Μζ÷ΤΓ±’β“ΜΦΌ…ηά¥ΒςΚΆ’β–©œύΜΞΟ§ΕήΒΡΥΒΖ®ΓΘΈ“Ο« Ι”ΟΡ‘Βγά¥Φλ―ι’β“ΜΦΌ…ηΘ§≤βΝΩ≤Έ”κ’Ώ‘Ύ1Ηω‘¬ΒΡ ±Φδάο÷¥––―Γ‘ώ–‘Ω’ΦδΉΔ“β»ΈΈώ ±¥ΧΦΛ”’ΖΔΒΡ ”ΨθΖ¥”ΠΓΘ‘Ύ―ΒΝΖ‘γΤΎΘ§Ω’ΦδΉΔ“β»ΈΈώΒΦ÷¬¥ΧΦΛ“ΐΤπΒΡ ”ΨθΖ¥”Π‘ω“φ‘ωΦ”ΓΘ‘ω“φ‘Ύ¥ΧΦΛΩΣ ΦΚσ‘Φ100msΡΎ≥ωœ÷Θ§“ΜΗωΜυ”Ύ–≈Κ≈Φλ≤βάμ¬έ(SDTΘ§ signal detection theory)ΒΡΕ®ΝΩΡΘ–Ά≥…ΙΠΒΊΫΪ’β÷÷‘ω“φΒς÷ΤΒΡΖυΕ»”κΉΔ“βΝΠœύΙΊΒΡ––ΈΣΗΡ…ΤΝΣœΒΤπά¥ΓΘ»ΜΕχΘ§‘Ύ≥ΛΤΎ―ΒΝΖ÷°ΚσΘ§’β÷÷‘γΤΎΒΡΉΔ“β‘ω“φœϊ≥ΐΝΥΘ§ΨΓΙή”κΉΔ“βΝΠœύΙΊΒΡ––ΈΣ»‘”– Β÷ –‘ΒΡΗΡ…ΤΓΘ“ρ¥ΥΘ§Μυ”ΎSDTΒΡΡΘ–Ά–η“ΣΫΒΒΆ‘κ“τΘ§“‘Ϋβ Ά¥ΧΦΛ”’ΖΔΒΡ ”ΨθΖ¥”ΠΚΆΉΔ“β––ΈΣΒςΫΎ÷°ΦδΒΡΝΣœΒΓΘ’β–©ΖΔœ÷±μΟςΘ§―ΒΝΖ¥”Ηυ±Ψ…œΗΡ±δΝΥΉΔ“βΗΡ±δ‘γΤΎΤΛ≤ψΖ¥”ΠΒΡΖΫ ΫΘ§Εχ’β–©‘γΤΎΤΛ÷ Ζ¥”Π «÷ß≥÷―Γ‘ώ–‘–≈œΔ¥ΠάμΒΡΓΘ¥ΥΆβΘ§’β–© ΐΨίΜΙ¥ΌΫχΝΥΩγ≤ΜΆ§Έο÷÷ΚΆΩγ≤…”Ο≤ΜΆ§––ΈΣ―ΒΝΖ÷ΤΕ»ΒΡ Β―ι≥Χ–ρΒΡΫαΙϊΉΣΜΜΓΘ
≤ΡΝœ”κΖΫΖ®
±Μ ‘
±Ψ―–ΨΩΙ≤’–ΡΦ23Οϊ ”ΝΠ’ΐ≥ΘΜρΫΟ’ΐΒΫ’ΐ≥Θ ”ΝΠΒΡΫΓΩΒ÷Ψ‘Η’ΏΓΘ≤…”Ο ΐΨίΆξ’ϊΒΡ12Οϊ≤Έ”κ’ΏΫχ––Ζ÷Έω(7Οϊ≈°–‘Θ§20-26ΥξΘ§ΨυΈΣ”“άϊ ÷)ΓΘ
¥ΧΦΛ”κ»ΈΈώ
Ι”ΟMATLABΚΆPsychophysics Toolbox (Αφ±Ψ3.0.8)‘ΎWindows XP…œ≥ œ÷¥ΧΦΛΓΘ≤Έ”κ’Ώ‘ΎΗτ“τΚΆΒγ¥≈ΤΝ±ΈΒΡΖΩΦδάοΘ§Ήχ‘ΎΨύάκCRTœ‘ ΨΤςΘ®Μ“…Ϊ±≥ΨΑ34.51cd/m2Θ§ΥΔ–¬¬ 60 HzΘ©60 cm¥ΠΓΘ
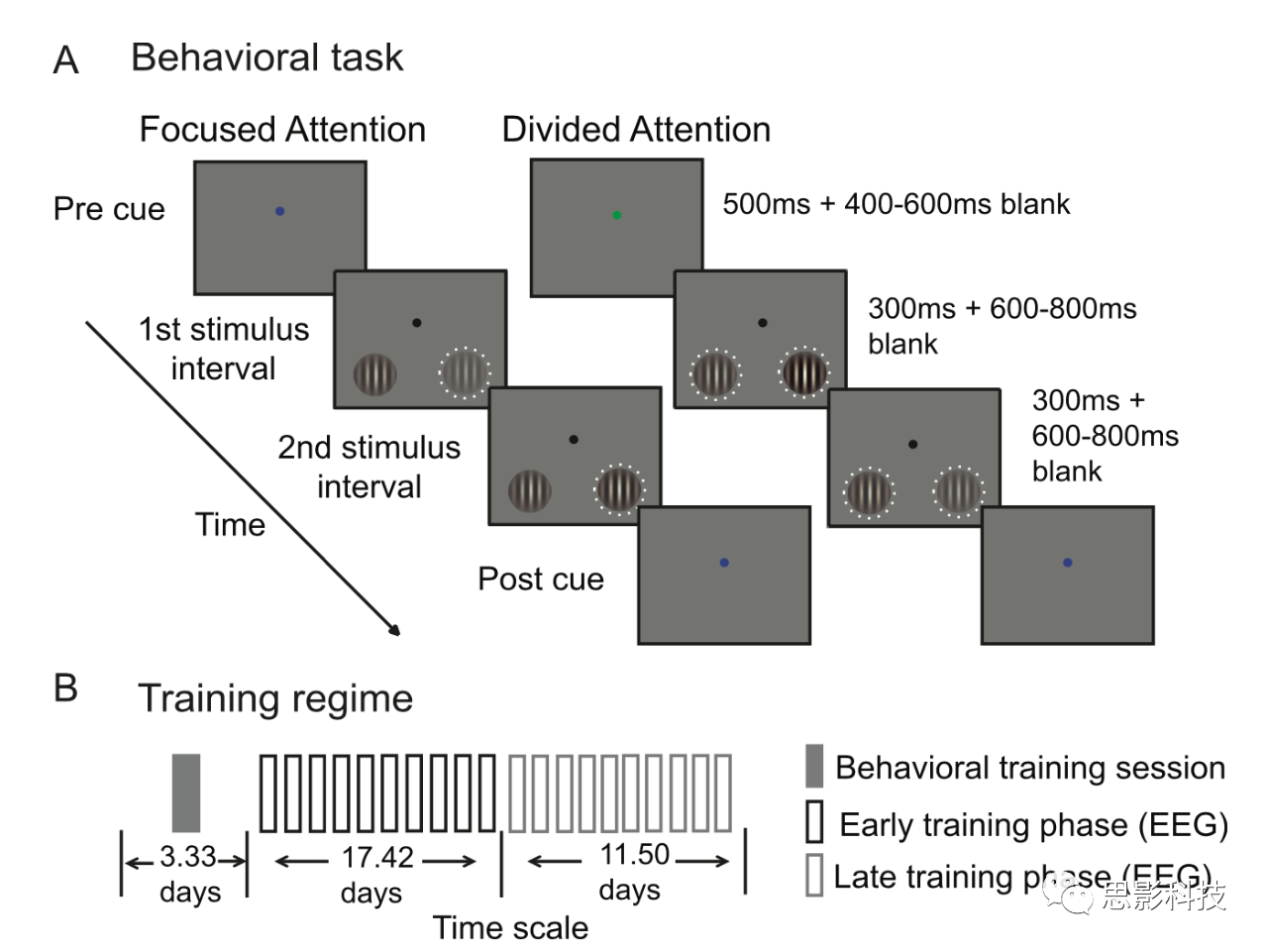
≤Έ”κ’Ώ÷¥––2IFCΘ®2-interval forced choiceΘ©Ε‘±»±φ±π»ΈΈώ(ΆΦ1A)ΓΘΟΩ“ΜΗω ‘¥ΈΩΣ Φ ±ΕΦ”–“ΜΗω≤ …ΪΒΡ≤ΌΉςΥΒΟςΓΘΚλ…ΪΧα ΨΉΔ“βΉσœ¬œσœόΘ§άΕ…ΪΧα ΨΉΔ“β”“œ¬œσœό(Φ·÷–ΉΔ“βΝΠ)ΘΜ¬Χ…ΪΧα Ψ±μ Ψ≤Έ”κ’Ώ”ΠΗΟΉΔ“βΝΫΗωœ¬œσœό(Ζ÷…ΔΉΔ“βΝΠ)ΓΘΦ·÷–ΉΔ“βΝΠœΏΥς «100%”––ßΒΡΘ§Φ¥Ήή «÷Η ΨΡΩ±ξΒΡΈΜ÷ΟΘ§ΕχΖ÷…ΔΉΔ“βΝΠœΏΥς±μΟςΘ§ΡΩ±ξ≥ωœ÷‘ΎΉσ≤ύœσœόΜρ”“≤ύœσœόΒΡΩ…Ρή–‘œύΒ»ΓΘ
precue≥ œ÷ΒΡ ±ΦδΈΣ500msΘ§»ΜΚσ «400-600msΒΡΩ’ΑΉ¥ΧΦΛΦδΗτ(ISI)ΓΘ÷°ΚσΝ§–χ≥ œ÷ΝΫΗωΒΡ¥ΧΦΛΘ§ΟΩΗω¥ΧΦΛΑϋΚ§ΈΜ”ΎΉσœ¬œσœόΚΆ”“œ¬œσœόΒΡ“ΜΕ‘’ΐœ“Gabor¥ΧΦΛ‘¥(Ω’ΦδΤΒ¬ ΈΣ1.04 cycles/degreeΘΜΗΏΥΙ¥Α±ξΉΦΤΪ≤νΈΣ1.9˚)ΓΘΟΩΕ‘¥ΧΦΛ≥ œ÷300msΘ§»ΜΚσ «600-800msΒΡISIΘ®Ω’ΑΉ¥ΧΦΛΦδΗτΘ©ΓΘΜυ”ΎGabor¥ΧΦΛΒΡ6ΗωΕ‘±»Ε»ΥφΜζ≥ωœ÷ΘΚ0%ΓΔ2.24%ΓΔ5.13%ΓΔ11.75%ΓΔ26.92%ΚΆ61.66%Θ®Ε‘±»Ε»«Ω»θ–ßΙϊΩ…ΦϊΆΦ3AΘ©ΓΘ‘ΎΟΩΗω ‘¥Έ÷–Θ§Ήσ”“GaborΒΡΖΫœρ «œύΆ§ΒΡΓΘΝΫΗω¥ΧΦΛ÷–ΒΡ“ΜΗω¥ΧΦΛΒΡΕ‘±»Ε» «¥”0%-61.66%Ε‘±»Ε»÷–Έ±ΥφΜζ≥ι»ΓΒΡΓΘΈ“Ο«ΫΪ’β–©Ε‘±»Ε»÷Β≥ΤΈΣΓΑΜυΓ±Ε‘±»Ε»ΓΘ‘ΎΝμ“ΜΗω¥ΧΦΛ÷–Θ§Έ“Ο«‘ΎΜυΕ‘±»Ε»÷Β…œ‘ωΦ”ΝΥ«αΈΔΒΡΕ‘±»Ε»‘ωΝΩ(ΠΛc)ΓΘ‘ΎΒΎΕΰ¥Έ¥ΧΦΛ÷°ΚσΘ§postcueΧα Ψ≤Έ”κ’ΏΉσ≤ύ¥ΧΦΛΘ®Κλ…ΪΧα ΨΘ©Μρ”“≤ύ¥ΧΦΛΘ®άΕ…ΪΧα ΨΘ©ΑϋΚ§’βΗωΕ‘±»Ε»‘ωΝΩΡΩ±ξΓΘ≤Έ”κ’Ώ–η±®Ηφ‘ωΝΩ «ΖΔ…ζ‘ΎΒΎ“ΜΗωΜρΒΎΕΰΗω¥ΧΦΛœ‘ Ψ÷–ΓΘΥϊΟ«±ΜΗφ÷Σ“Σ”≈œ»ΩΦ¬«ΉΦ»Ζ–‘Θ§Εχ«“ΟΜ”–ΜΊΗ¥ΒΡ ±Φδœό÷ΤΓΘ
‘ΎΦ·÷–ΉΔ“βΧθΦΰœ¬Θ§”–Χα ΨΈΜ÷ΟΒΡΝΫΗωΝ§–χ¥ΧΦΛΉή «“‘≤ΜΆ§ΒΡΕ‘±»Ε»≥ œ÷ΓΘΟΜ”–Χα ΨΈΜ÷ΟΒΡΝΫΗωΝ§–χΒΡ¥ΧΦΛΉή «“‘œύΆ§ΒΡΕ‘±»Ε»≥ œ÷ΓΘΈ“Ο«ΫΪ‘ΎœΏΥςΈΜ÷Ο÷–≥ œ÷ΒΡ¥ΧΦΛ≥ΤΈΣΓΑΦ·÷–ΡΩ±ξΓ±¥ΧΦΛΘ§ΕχΫΪΖ«œΏΥςΈΜ÷Ο÷–≥ œ÷ΒΡ¥ΧΦΛ≥ΤΈΣΓΑΦ·÷–Ζ«ΡΩ±ξΓ±¥ΧΦΛΓΘ‘ΎΖ÷…ΔΉΔ“βΝΠΒΡ«ιΩωœ¬Θ§ΝΫΗωΈΜ÷ΟΕΦ”–Ω…ΡήΑϋΚ§Ε‘±»Ε»ΒΡ±δΜ·Θ§≤ζ…ζΓΑΖ÷…ΔΡΩ±ξΓ±ΚΆΓΑΖ÷…ΔΖ«ΡΩ±ξΓ±¥ΧΦΛΓΘ÷ς“ΣΒΡ“άάΒ–‘≤βΝΩ «ΟΩΗωΜυΕ‘±»Ε»ΒΡΕ‘±»Ε»±δΜ·(Φ¥Ε‘±»Ε»ψ–÷ΒΜρΠΛc)–η“Σ¥οΒΫ76%ΒΡΉΦ»ΖΕ»(Η–÷ΣΝιΟτΕ»Μρd'‘ΦΈΣ1)ΓΘΒ±”κEEGΆ§≤ΫΦ«¬ΦœύΫαΚœ ±Θ§’β÷÷–ΡάμΈοάμΖΫΖ® ΙΈ“Ο«ΡήΙΜΆΤΒΦ≥ω–Ράμ≤βΝΩΚΆ…ώΨ≠≤βΝΩΖ¥”ΠΚ· ΐΘ§’β―υΈ“Ο«ΨΆΩ…“‘÷±Ϋ”ΫΪ…ώΨ≠Ζ¥”Π÷–ΒΡΉΔ“βΝΠΒςΫΎ”κ––ΈΣ÷–ΒΡΉΔ“βΝΠΒςΫΎΝΣœΒΤπά¥ΓΘ

ΆΦ1. Β―ι»ΈΈώ…ηΦΤΓΘ
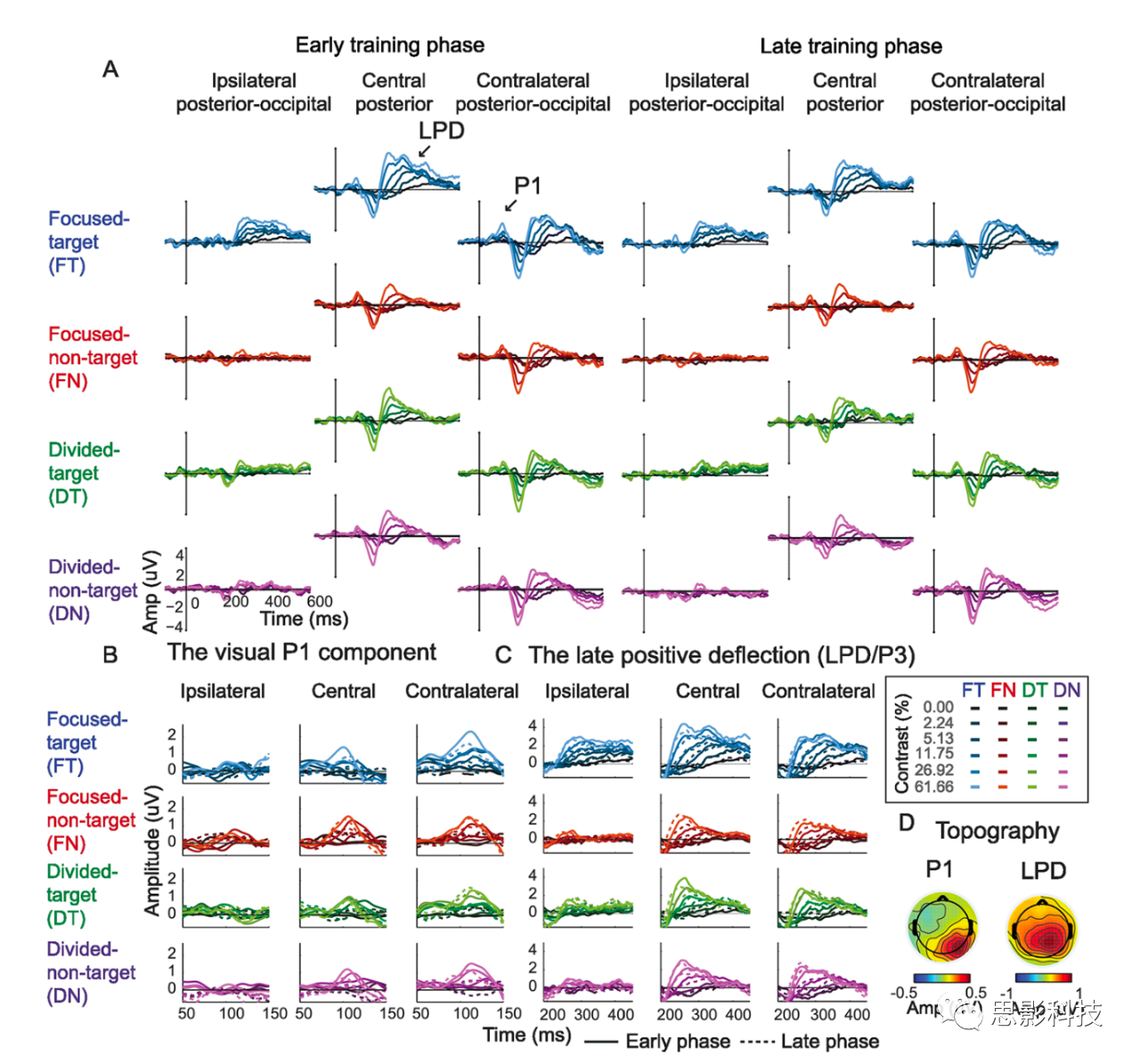
(A)–η“ΣΦ·÷–ΜρΖ÷…ΔΩ’ΦδΉΔ“βΒΡ2IFCΕ‘±»±φ±π»ΈΈώΓΘ
(B)―ΒΝΖΖΫΑΗΘΚ12Οϊ≤Έ”κ’ΏΆξ≥…ΝΥ1¥Έ––ΈΣ―ΒΝΖΘ§»ΜΚσ‘ΎΤΫΨυ32.25ΧλΡΎΆξ≥…ΝΥ20¥ΈΆ§ ±Ϋχ––ΒΡ––ΈΣ≤β ‘ΚΆEEGΦ«¬ΦΓΘΈΣΝΥΫχ––÷ς“ΣΖ÷ΈωΘ§Έ“Ο«ΫΪ«Α10¥Έ≤β ‘Ε®“εΈΣ―ΒΝΖΒΡΓΑ‘γΤΎΫΉΕΈΓ±Θ§ΫΪΚσ10¥Έ≤β ‘Ε®“εΈΣΓΑΚσΤΎΫΉΕΈΓ±ΓΘ
‘ΎΒΎ“ΜΧλΘ§≤Έ”κ’Ώ≤ΈΦ”ΝΥ‘Φ2.5–Γ ±ΒΡ––ΈΣ―ΒΝΖΘ§Τδ÷–”Π”ΟΫΉΧί≥Χ–ρά¥ΙάΦΤΟΩ÷÷ΉΔ“βΧθΦΰΚΆΟΩ÷÷Ε‘±»Ε»Υ°ΤΫΒΡΕ‘±»Ε»±φ±πψ–÷ΒΓΘ’β–©ψ–÷ΒΥφΚσ‘ΎΒΎ“Μ¥ΈEEG≤β ‘÷– Ι”ΟΓΘ≤Έ”κ’ΏΆξ≥…20¥ΈEEG≤β ‘(ΟΩ÷ή2-3ΧλΘ§ΟΩΧλ1-2¥ΈΘ§Υυ”–≤Έ”κ’ΏΤΫΨυ‘Ύ12.08ΧλΡΎΆξ≥…2Ηω¥Έ)ΓΘΟΩ¥ΈEEG≤β ‘ΑϋΚ§8ΗωΡΘΩιΘ§Ι≤288¥ΈΘ§Υυ”– Β―ιΧθΦΰΨυ¥οΒΫΤΫΚβΘΚ2(ΉΔ“βΧθΦΰΘΚΦ·÷–ΓΔΖ÷…Δ)ΓΝ2(ΡΩ±ξΈΜ÷ΟΘΚΉσΓΔ”“)ΓΝ2(ΡΩ±ξ¥ΧΦΛΘΚΒΎ“ΜΓΔΒΎΕΰ)ΓΝ6(ΡΩ±ξΕ‘±»Ε»Υ°ΤΫ)ΓΝ6(Ζ«ΡΩ±ξΕ‘±»Ε»Υ°ΤΫ)ΓΘ
‘ΎΟΩ¥ΈEEG≤β ‘ΚσΘ§Βς’ϊΟΩΗωΉΔ“βΧθΦΰΚΆΟΩΗωΡΩ±ξΜυΕ‘±»Ε»ΒΡΕ‘±»Ε»ψ–÷Β(ΠΛc)Θ§“‘±ψ‘ΎΥυ”– Β―ιΧθΦΰœ¬ΫΪΉΦ»Ζ¬ ±Θ≥÷‘Ύ~76%(d'=~1)ΓΘ‘Ύ12Οϊ≤Έ”κ’Ώ÷–Θ§ΒΎ“Μ¥Έ––ΈΣ―ΒΝΖ”κΒΎ“Μ¥ΈEEG≤β ‘÷°ΦδΓΔΒΎ“Μ¥Έ”κΒΎ11¥ΈEEG≤β ‘÷°Φδ(‘γΤΎ―ΒΝΖΫΉΕΈ)“‘ΦΑΒΎ11¥Έ”κΉνΚσ“Μ¥ΈEEG≤β ‘÷°ΦδΒΡΤΫΨυ ±ΦδΖ÷±πΈΣ3.33Γά0.66ΧλΓΔ17.41Γά2.31ΧλΚΆ11.50Γά1.12ΧλΘ®meanΓάSEMΘ© (ΆΦ1B)ΓΘ
––ΈΣΖ÷Έω
Υυ”–ΉΔ“βΧθΦΰ(Φ·÷–ΉΔ“βΚΆΖ÷…ΔΉΔ“β)ΓΔ―ΒΝΖΫΉΕΈ(‘γΤΎΚΆΚσΤΎ)ΚΆ¥ΧΦΛΕ‘±»(0%~61.66%)ΒΡΕ‘±»±φ±πψ–÷ΒΒΡΉΦ»Ζ¬ ΈΣ76%(d'=1)ΓΘ‘Ύ÷ς“ΣΒΡ––ΈΣΖ÷Έω÷–Θ§Έ“Ο«ΫΪ ΐΨίΖ÷ΈΣΝΫΗω―ßœΑΫΉΕΈ(‘γΤΎΒΡ«Α10ΗωEEG≤β ‘ΚΆΚσΤΎΒΡΚσ10ΗωEEG≤β ‘)Θ§“‘”κ÷ς“ΣΒΡEEGΖ÷ΈωœύΤΞ≈δΓΘ≤…”Ο»ΐ“ρΥΊ÷ΊΗ¥≤βΝΩΖΫ≤νΖ÷ΈωΖΫΖ®Θ§ΫαΚœΉΔ“βΧθΦΰΓΔ―ΒΝΖΫΉΕΈΚΆ¥ΧΦΛΕ‘±»Ε»Β»±Μ ‘ΡΎ“ρΥΊΘ§Φλ―ι’β–©“ρΥΊΒΡ÷ς–ß”ΠΦΑΫΜΜΞΉς”ΟΓΘ»ΜΚσ Ι”ΟPost-hoc≈δΕ‘tΦλ―ιά¥Φλ―ιΉΔ“β–ß”ΠΚΆ―ßœΑ–ß”ΠΕ‘ΟΩΗωΕ‘±»Ε»Υ°ΤΫΒΡΕ‘±»Ε»±φ±πψ–÷Β ΐΨίΒΡ”ΑœλΘ§≤Δ”ΟΠΝ÷ΒΈΣ0.05ΒΡFDRΖ®Ε‘Εύ÷Ί±»ΫœΫχ–––Θ’ΐΓΘΈ“Ο«‘Ύ’βάο Ι”ΟΝΥΒΞΈ≤Ά≥ΦΤ ΐΨίΘ§ΦΌ…ηΥφΉ≈ΉΔ“βΝΠΚΆ―ΒΝΖΒΡ‘ωΦ”Θ§––ΈΣ±μœ÷”ΠΗΟΜα”–ΥυΗΡ…ΤΓΘ
ΈΣΝΥ‘ΎΗϋΨΪœΗΒΡ ±Φδ≥ΏΕ»…œΦλ―ι―ΒΝΖ–ßΙϊΘ§Έ“Ο«ΜΙΫχ––ΝΥ÷π»’ΒΡ––ΈΣΖ÷ΈωΘ§ΫΪΕ‘±»±φ±πψ–÷ΒΖ÷ΈΣ10Ηω ±ΦδΕΈ(ΟΩΗω ±ΦδΕΈ2¥ΈΡ‘Βγ ΐΨί)ΓΘ“‘ ±ΦδΈΣ±Μ ‘ΡΎ“ρΥΊΘ§≤…”Ο÷ΊΗ¥≤βΝΩΒΡΒΞ“ρΥΊΖΫ≤νΖ÷ΈωΖΫΖ®Θ§Ζ÷±πΩΦ≤λΝΥ‘ΎΦ·÷–ΉΔ“βΚΆΖ÷…ΔΉΔ“βΧθΦΰœ¬Θ§―ΒΝΖΕ‘≤ΜΆ§Ε‘±»Ε»Υ°ΤΫ¥ΧΦΛΒΡΕ‘±»Ε»±φ±πψ–÷ΒΒΡ”ΑœλΘ§ΫαΙϊ±μΟςΘ§―ΒΝΖΕ‘≤ΜΆ§Ε‘±»Ε»Υ°ΤΫ¥ΧΦΛΒΡΕ‘±»Ε»±φ±πψ–÷Β”–≤ΜΆ§≥ΧΕ»ΒΡ”ΑœλΓΘΈΣΝΥΨ≤Χ§ΒΊΤάΙά±φ±πψ–÷ΒΒΡ÷π»’±δΜ·Θ§Ϋχ––ΝΥpost hoc≈δΕ‘tΦλ―ιΘ§“‘Φλ≤ιΒΎ“ΜΧλΚΆΒΎΕΰΧλ÷°Φδ“‘ΦΑΒΎΕΰΧλΚΆΤδ”ύΧλ ΐ÷°ΦδΒΡ ΐΨίψ–÷ΒΒΡ≤ν“λΓΘ
EEG ¥Πάμ”κΖ÷Έω
Έ“Ο«”Ο64+8Ά®ΒάΒΡBiosemi ActiveTwoœΒΆ≥Θ®ΨΏΧεΫι…ή«κΒψΜςΘΚBIOSEMIΡ‘ΒγœΒΆ≥Ϋι…ήΘ©“‘512 HzΒΡ≤…―υ¬ Φ«¬ΦΡ‘Βγ ΐΨίΓΘΥυ”––≈Κ≈”κCMS-DRLΜυΉΦΒΡΤΪ“ΤΕΦ±Θ≥÷‘Ύ20 uV“‘œ¬ΓΘΈ“Ο« Ι”ΟEEGlab11.0.3.1bΚΆΉ‘Ε®“εΒΡMATLABΫ≈±ΨΕ‘EEG ΐΨίΫχ––‘Λ¥ΠάμΓΘ
1Θ© Ι”ΟΥΪ≤ύ»ιΆΜΕ‘ ΐΨίΫχ––÷Ί≤ΈΩΦ
2Θ© Ι”ΟButterworth¬Υ≤®Τς(»ΐΫΉ)Ε‘ ΐΨίΫχ––ΘΚΗΏΆ®0.25 HzΚΆΒΆΆ®55 Hz¬Υ≤®ΓΘ
3Θ©“‘¥ΧΦΛΩΣ ΦΒΡ-500ms~3500msΕ‘ ΐΨίΫχ––Ζ÷ΕΈΓΘ
4Θ© Ι”ΟΕάΝΔ≥…Ζ÷Ζ÷Έω(ICA)»Ξ≥ΐ―έΒγ≥…Ζ÷ΓΘ
5Θ©»Ξ≥ΐΦΓΒγΒ»œύΙΊΈ±ΦΘΓΘ
6Θ©Ε‘ ΐΨίΫχ––ΜυœΏ–Θ’ΐΘΚ0-200msΓΘ
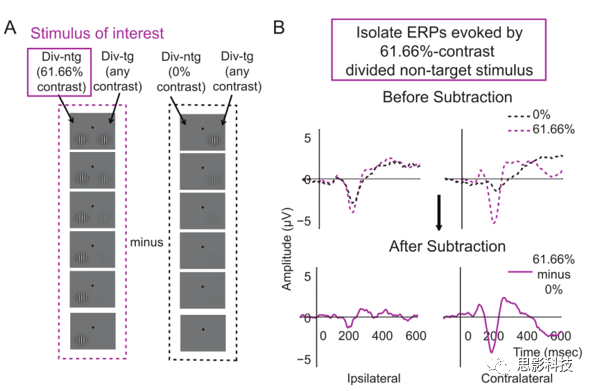
ΫΪ‘Λ¥ΠάμΚσΒΡΡ‘Βγ ΐΨίΫχ––Ζ÷άύΘΚ2ΗωΉΔ“βΧθΦΰ(Φ·÷–ΚΆΖ÷…ΔΉΔ“β)ΓΝ2Ηω¥ΧΦΛάύ–Ά(ΡΩ±ξΚΆΖ«ΡΩ±ξ)ΓΝ2Ηω―ΒΝΖΫΉΕΈ(‘γΤΎΚΆΆμΤΎ)ΓΝ6Ηω¥ΧΦΛΕ‘±»Ε»Υ°ΤΫx 2Ηω¥ΧΦΛΦδΗτ(ΒΎ“ΜΚΆΒΎΕΰΗω)x 2Ηω¥ΧΦΛΈΜ÷Ο(ΉσΚΆ”“)ΓΘ»ΜΚσΘ§Ά®Ιΐ…œ ωΖ÷άύΒΡΡ‘Βγ ΐΨίΖ÷±πά¥ΦΤΥψ¥ΧΦΛΥχΕ®ΒΡ ¬ΦΰœύΙΊΒγΈΜΓΘΈΣΝΥΧα»Γ”…Η––Υ»ΛΒΡ¥ΧΦΛ“ΐΤπΒΡERPs(Φ¥Θ§Φθ»Ξ”…”κΗ––Υ»ΛΒΡ¥ΧΦΛ≈δΕ‘ΒΡ¥ΧΦΛ“ΐΤπΒΡΖ¥”Π)Θ§≤Δ¥”œΏΥς÷–Ήν¥σœόΕ»ΒΊΦθ…Ό»ΈΚΈ‘ΛΤΎ–ß”ΠΒΡΜλœΐΘ§Έ“Ο«¥”Υυ”–ΤδΥϊΧθΦΰœ¬ΒΡERPs÷–Φθ»Ξ”…0%Ε‘±»Ε»¥ΧΦΛ(Φ¥Θ§Β±œύΕ‘”ΎΗχΕ®EEGΒγΦΪΒΡΕ‘≤ύ ”“Α÷–ΟΜ”–¥ΧΦΛ ±)”’ΖΔΒΡERPs (ΆΦ3)ΓΘ’β”–÷ζ”ΎΖ÷άκ”κΕ‘≤ύ¥ΧΦΛ≥ œ÷ΧΊ±πœύΙΊΒΡERPΓΘΫΪΗ––Υ»ΛΒΡ¥ΧΦΛ(Φ·÷–ΡΩ±ξΓΔΦ·÷–Ζ«ΡΩ±ξΓΔΖ÷…ΔΡΩ±ξΚΆΖ÷…ΔΖ«ΡΩ±ξΧθΦΰ)“ΐΤπΒΡCRFs(contrast response functions,Ε‘±»œλ”ΠΚ· ΐ)”κΆ§ ±≥ œ÷‘Ύœ‘ ΨΤςΝμ“Μ≤ύΒΡ¥ΧΦΛ(άΐ»γΘ§»γΙϊΗ––Υ»ΛΒΡ¥ΧΦΛ «Φ·÷–ΡΩ±ξΘ§‘ρ”κΤδ≈δΕ‘ΒΡ¥ΧΦΛΫΪ «Φ·÷–Ζ«ΡΩ±ξ)Ζ÷άκ «÷ΝΙΊ÷Ί“ΣΒΡΓΘ“ρ¥ΥΘ§Φθ»ΞΆ§≤ύ¥ΧΦΛ“ΐΤπΒΡΖ¥”Π”–÷ζ”ΎΧαΗΏERPΖ¥”ΠΒΡΩ’Φδ―Γ‘ώ–‘ΓΘ¥ΥΆβΘ§‘Ύ≤ΜΦθ»Ξ0%Ε‘±»Ε»ERPΒΡ«ιΩωœ¬Θ§»ΈΚΈΉΔ“βΒςΫΎΕΦ”–Ω…Ρή±ΜœΏΥςœύΙΊΒΡΖ«Ω’Φδ―Γ‘ώ–‘‘ΛΤΎΖ¥”ΠΥυΜλœΐΘ§Εχ≤Μ «¥ΧΦΛ”’ΖΔΖ¥”ΠΒΡΉΔ“βΒςΫΎ(Φ¥ΜΫ–―ΒΡ±δΜ·Θ§Εχ≤Μ «―Γ‘ώ–‘Ω’ΦδΉΔ“βΒΡ±δΜ·)ΓΘΈ“Ο«“≤ΑϋΚ§ΝΥΟΜ”–ΜυœΏΦθΖ®ΒΡΫαΙϊΘ§“‘―ι÷ΛΗΟΖΫΖ®ΒΡœΏ–‘ΦΌ…ηΓΘ
ΦΤΥψ¥ΧΦΛΚσ80-130ms ”ΨθP1≥…Ζ÷‘ΎΕ‘≤ύ-Κσ≤ΩΒγΦΪ…œΒΡΤΫΨυ’ώΖυΘ§Τδ÷–P1‘ΎΥυ”– Β―ιΧθΦΰœ¬ΒΡΤΫΨυ’ώΖυΉνΗΏΒγΦΪΘΚΉσΑκ«ρΈΣPO7ΓΔP5ΚΆP7Θ§”“Ακ«ρΈΣPO8ΓΔP6ΚΆP8ΓΘ ±Φδ¥ΑΩΎΒΡ―Γ‘ώΜυ”Ύ«Α»ΥΕ‘ ”ΨθΉΔ“βΒΡERP―–ΨΩΘ§“‘50msΒΡ¥ΑΩΎ¥σ–ΓΉςΈΣ±ξΉΦΓΘ»ΜΚσΫΪP1ΖυΕ»Ψυ÷ΒΜφ÷ΤΈΣ¥ΧΦΛΕ‘±»Ε»ΒΡΚ· ΐΘ§Ζ÷±πΈΣΟΩΗωΉΔ“βΧθΦΰΓΔΟΩΗω¥ΧΦΛάύ–ΆΚΆΟΩΗω―ΒΝΖΫΉΕΈΧαΙ©Μυ”ΎP1ΒΡCRFsΓΘ‘ΎCRFsΒΡx÷α…œΘ§Φ·÷–ΚΆΖ÷…ΔΒΡΖ«ΡΩ±ξ¥ΧΦΛΕ‘±»Ε»Ζ÷±πΙΧΕ®ΈΣ0%ΓΔ2.24%ΓΔ5.13%ΓΔ11.75%ΓΔ26.92%ΚΆ61.66%ΓΘ»ΜΕχΘ§”…”ΎΡΩ±ξ–ρΝ–ΑϋΚ§Μυ¥ΧΦΛΚΆ‘ωΝΩ¥ΧΦΛΘ§Έ“Ο« Ι”ΟΝΫΗω¥ΧΦΛ÷°ΦδΒΡΤΫΨυΕ‘±»Ε»÷Βά¥Μφ÷ΤΦ·÷–ΡΩ±ξΚΆΖ÷…ΔΡΩ±ξΧθΦΰœ¬ΒΡCRFΓΘ Ι”ΟLoftusΚΆMassonΖΫΖ®ΦΤΥψΟΩΗωΕ‘±»Ε»Υ°ΤΫΒΡ ΐΨίΒΡ≤Έ”κ’ΏΡΎSEM(Έσ≤νΘ©ΓΘΫ”œ¬ά¥Θ§Ά®ΙΐΕ‘≤Έ”κ’ΏΫχ––÷Ί≤…―υΘ§Ε‘P1 ΐΨίΫχ–– bootstrappedΘ§≤ΔΫχ––ΧφΜΜΘ§¥Έ ΐΈΣ10000¥ΈΓΘ‘ΎΟΩ¥ΈbootstrappedΒϋ¥ζ÷–Θ§ΟΩΗωΉΔ“βΧθΦΰΓΔ¥ΧΦΛάύ–ΆΚΆ―ΒΝΖΫΉΕΈΒΡCRF ΐΨί”ΟNaka-RushtonΖΫ≥ΧΡβΚœΘΚ

Τδ÷–Θ§R(c) «ΉςΈΣ¥ΧΦΛΕ‘±»Κ· ΐΒΡP1ΖυΕ»Θ§Gr «ΩΊ÷ΤCRFΒΡ¥Ι÷±“ΤΈΜΒΡ≥ΥΖ®œλ”Π‘ω“φ“ρΉ”Θ§Gc «ΩΊ÷ΤCRFΒΡΥ°ΤΫ“ΤΈΜΒΡΕ‘±»Ε»‘ω“φ“ρΉ”Θ§b «œλ”ΠΜυœΏΤΪ“Τ,q «ΩΊ÷ΤCRF…œ…ΐ≤Δ¥οΒΫΫΞΫϋœΏΒΡΥΌΕ»ΒΡ÷Η ΐΓΘΦχ”ΎΙΐ»ΞΩ’ΦδΉΔ“βΒΡΡ‘Βγ―–ΨΩ“Μ÷±±®ΗφΜυ”ΎEEGΒΡCRFΖ¥”ΠΜυœΏΟΜ”–±δΜ·Θ§‘Ύ±Ψ―–ΨΩ÷–Θ§¥”Υυ”– ‘¥Έ÷–Φθ»ΞΕ‘0%Ε‘±»¥ΧΦΛΒΡ”’ΖΔΖ¥”Π( ΙΒΟERP‘Ύ0%Ε‘±» ‘―ι÷–≥÷ΤΫ)bΙΧΕ®ΈΣΥυ”– Β―ιΧθΦΰœ¬Ήν–ΓΖυΕ»ΒΡΤΫΨυ÷ΒΓΘ»ΜΚσΘ§Έ“Ο« Ι”ΟΉν–ΓΕΰ≥ΥΈσ≤νΙάΦΤΖΫΖ®(MATLAB÷–ΒΡfminsearchΚ· ΐ)ά¥ΙάΦΤΉν¥σœλ”Π(100%Ε‘±»Ε»Φθ»ΞΜυœΏ ±ΒΡœλ”Π)ΓΔΑκΉν¥σΕ‘±»Ε»(œλ”Π¥οΒΫΉν¥σ“ΜΑκ ±ΒΡΕ‘±»Ε»)ΚΆ÷Η ΐ(q)≤Έ ΐΓΘ”…”Ύ‘Ύ–μΕύ Β―ιΧθΦΰœ¬Θ§CRF‘ΎΉν¥σΕ‘±»Ε»Υ°ΤΫ(100%)œ¬ΟΜ”–Άξ»Ϊ±ΞΚΆΘ§Έ“Ο«œό÷ΤΝΥΡβΚœΙΐ≥ΧΘ§“‘±ψΉν¥σœλ”Π÷Β≤ΜΡή≥§Ιΐ61.66%Ε‘±»Ε»÷Β(¥ΧΦΛΦ·÷–ΉνΗΏΕ‘±»Ε»)œ¬ΒΡΖ¥”ΠΒΡ1.5±ΕΓΘGrΚΆGc ήΒΫ‘Φ χΘ§“ρ¥ΥΥϋΟ«≤ΜΡήΖ÷±π–Γ”Ύ0ΚΆ1ΓΘ÷Η ΐq“≤±Μœό÷Τ‘Ύ-10ΒΫ10ΒΡΖΕΈßΡΎΓΘΈ“Ο«Ζ÷±π Ι”Ο30%ΒΡΕ‘±»Ε»÷Β(‘ΦΈΣ61.66%Ε‘±»Ε»ΒΡ“ΜΑκ)ΉςΈΣGcΒΡ≥θ Φ÷÷Ή”÷ΒΘ§ Ι”ΟΉν¥σΚΆΉν–Γœλ”Π÷°ΦδΒΡ≤ν÷ΒΉςΈΣGrΒΡ÷÷Ή”÷ΒΘ§ Ι”Ο1ΚΆ5ΉςΈΣ÷Η ΐqΜυ”ΎP1ΚΆLPDΡβΚœCRF ±ΒΡ÷÷Ή”÷ΒΘ®LPDΦϊœ¬ΖΫΘ©ΓΘ÷Η ΐqΒΡ≥θ Φ÷÷Ή”÷»ù‘Μυ”Ύœ»«Α―–ΨΩΒΡΙάΦΤ÷ΒΓΘ
Ε‘”ΎΟΩΗω―ΒΝΖΫΉΕΈΘ§Έ“Ο« Ήœ»≤β ‘ΝΥΉΔ“βΧα Ψ(Χα Ψ”κΈόΧα Ψ)Ε‘Μυ”ΎP1ΒΡCRF‘ΎΦ·÷–ΉΔ“βΧθΦΰœ¬ΒΡΉν¥σΖ¥”ΠΒΡ”ΑœλΓΘΈΣ¥ΥΘ§Έ“Ο«ΦΤΥψΦ·÷–ΡΩ±ξ(Φ¥Θ§ΉΔ“β)ΚΆΦ·÷–Ζ«ΡΩ±ξ(Φ¥Θ§Ζ«ΉΔ“β)ΧθΦΰ÷–ΒΡΙάΦΤΡβΚœ≤Έ ΐ÷°ΦδΒΡ≤ν÷ΒΒΡbootstrappedΖ÷≤ΦΘ§≤ΔΦΤΥψΗΟΖ÷≤ΦΒΡΈ≤≤Ω÷–¥σ”ΎΜρ–Γ”Ύ0ΒΡ÷ΒΒΡΑΌΖ÷±»(ΥΪΈ≤ΈΣ±Θ Ί)ΓΘΫ”œ¬ά¥Θ§Έ“Ο«Ά®ΙΐΦΤΥψΦ·÷–ΡΩ±ξΚΆΖ÷…ΔΉΔ“βΧθΦΰœ¬ΙάΦΤΒΡΡβΚœ≤Έ ΐ÷°≤νΒΡbootstrappedΖ÷≤ΦΘ§Φλ―ιΝΥΉΔ“βœΏΥςάύ–Ά(Φ·÷–ΉΔ“β”κΖ÷…ΔΉΔ“β)Ε‘Μυ”ΎP1ΒΡCRFΉν¥σΖ¥”ΠΒΡ”ΑœλΓΘ»ΜΚσΘ§Έ“Ο«ΦΤΥψΗΟΖ÷≤ΦΒΡΈ≤≤Ω÷–¥σ”ΎΜρ–Γ”Ύ0(ΥΪΈ≤)ΒΡ÷ΒΒΡΑΌΖ÷±»ΓΘΉΔ“βΘ§‘ΎΖ÷…ΔΉΔ“βΝΠΒΡ«ιΩωœ¬Θ§Έ“Ο«Κœ≤ΔΝΥΖ÷…ΔΡΩ±ξΚΆΖ÷…ΔΖ«ΡΩ±ξΧθΦΰ÷°ΦδΒΡ ΐΨίΘ§“ρΈΣ”κ’βΝΫΗωΧθΦΰœύΙΊΒΡP1ΖυΕ»ΟΜ”–≤ν“λ(≤ΈΦϊΆΦ5C)ΓΘΕ‘”ΎP1ΒΡΉν¥σΖ¥”ΠΘ§Έ“Ο«‘Ύ―ΒΝΖ‘γΤΎΙέ≤λΒΫΝΥ“ΜΗωrobustΒΡΉΔ“β‘ω“φΒς÷ΤΘ§ΒΪ’β÷÷‘ω“φΒς÷Τ‘Ύ―ΒΝΖΚσœϊ ßΝΥ(ΆΦ5ΚΆΆΦ6)ΓΘΈ“Ο«Ά®ΙΐΦλ―ι―ΒΝΖ(‘γΤΎΚΆΚσΤΎ)ΚΆΉΔ“βœΏΥςάύ–Ά(Φ·÷–ΚΆΖ÷…ΔΉΔ“β)÷°ΦδΒΡΫΜΜΞΉς”Οά¥Ε‘’β“ΜΙέ≤λΫαΙϊΫχ––Ά≥ΦΤΤάΙάΘ§≤Δ±»ΫœΝΥ‘γΤΎ―ΒΝΖΫΉΕΈΦ·÷–ΡΩ±ξΚΆΖ÷…ΔΉΔ“βΧθΦΰ÷°ΦδΒΡ≤ν“λΖ÷≤ΦΚΆ―ΒΝΖΚσΤΎΫΉΕΈ≤ν“λΒΡΖ÷≤ΦΓΘ»ΜΚσΘ§Έ“Ο«ΤάΙάΝΥΦ·÷–ΡΩ±ξΓΔΦ·÷–Ζ«ΡΩ±ξΚΆΖ÷…ΔΉΔ“βΧθΦΰœ¬ΒΡΉν¥σœλ”Π÷Β «Ζώ‘Ύ―ΒΝΖΫΉΕΈΖΔ…ζΝΥ±δΜ·ΓΘ»ΜΚσΕ‘”κΜυ”ΎP1ΒΡCRFœύΙΊΒΡΑκΉν¥σΕ‘±»Ε»÷ΒΫχ––œύΆ§ΒΡΖ÷ΈωΓΘ
Ε‘”ΎLPD≥…Ζ÷Θ§ΦΤΥψ¥ΧΦΛΚσ230-380ms¥©ΙΐΚσ≤ΩΚΆ’μΚσ≤ΩΒγΦΪ(P5Θ§P7Θ§PO7Θ§P1Θ§PzΘ§P2Θ§P6Θ§P8ΚΆPO8)ΒΡΤΫΨυ’ώΖυΓΘΗΟΖ÷Έω¥ΑΩΎΚΆΒγΦΪ «Μυ”ΎLPD’ώΖυΒΡΙψΖΚΦΛΜνΕχ―Γ‘ώΒΡΘ§≤Δ«“‘ΎΥυ”– Β―ιΧθΦΰΚΆ¥ΧΦΛΕ‘±»Ε»Υ°ΤΫ…œ»ΓΤΫΨυ÷ΒΓΘΜΙ“ΣΉΔ“βΘ§―Γ‘ώP1ΚΆLPD≥…Ζ÷ΒΡΖ÷Έω¥ΑΩΎ «ΈΣΝΥΫΪ≥ωœ÷‘Ύ¥ΧΦΛΚσ‘Φ150-200msΒΡN1ΗΚ≥…Ζ÷ΒΡ‘κ“τΫΒ÷ΝΉνΒΆ(≤ΈΦϊΆΦ4BΚΆ4C÷–P1ΚΆLPD≥…Ζ÷ΒΡΖ≈¥σERPΙλΦΘΘ§Τδ÷–‘Ύ’β–©¥ΑΩΎ…œΙέ≤λΒΫΉν–ΓΒΡΗΚΒγΈΜ)ΓΘΕ‘LPD ΐΨί“≤Ϋχ––ΝΥ…œ ωœύΆ§ΒΡbootstrappedΓΔΡβΚœΚΆΆ≥ΦΤΖ÷ΈωΓΘ
ΈΣΝΥΫχ––±»ΫœΘ§Έ“Ο«ΜΙΖ÷ΈωΝΥΟΜ”–Φθ»ΞΜυœΏΒΡP1ΚΆLPD ΐΨί(≤ΈΦϊΆΦ6ΚΆ11÷–ΒΡΫαΙϊ)ΓΘ Ήœ»Θ§Έ“Ο«‘Ύ”κ…œ ωœύΆ§ΒΡΒγΦΪΚΆœύΆ§ΒΡ ±Φδ¥ΑΩΎ÷–ΜώΒΟP1ΚΆLPD≥…Ζ÷Θ§≤ΔΗυΨίP1ΚΆLPDΒΡΤΫΨυΖυΕ»Μφ÷ΤCRFΓΘΒΪ «Θ§ΟΜ”– Β ©…œ ωΜυœΏΦθΖ®ΓΘΫ”œ¬ά¥Θ§Έ“Ο«ΡβΚœNaka-RushtonΖΫ≥Χ(Β» Ϋ1)ά¥±μ’ςCRFΘ§ΒΪ’β“Μ¥ΈΈ“Ο«Αϋά®ΝΥ“ΜΗωΕνΆβΒΡΉ‘”…≤Έ ΐά¥Ϋβ Ά≤ΜΆ§ΧθΦΰ÷°ΦδΒΡΜυœΏ≤ν“λΓΘ
»ΜΚσΘ§Έ“Ο«Ϋχ––«ΕΧΉΡΘ–Ά±»ΫœΘ§“‘ΤάΙά‘ –μΜυœΏ≤Έ ΐΉ‘”…ΗΡ±δΒΡΡΘ–Ά(ΈόΜυœΏΡΘ–Ά)ΚΆ‘ΎΥυ”– Β―ιΧθΦΰœ¬ΙΧΕ®ΜυœΏ≤Έ ΐΒΡΡΘ–ΆΘ®ΜυœΏΙΧΕ®ΡΘ–ΆΘ©÷°ΦδΒΡΡβΚœ”≈Ε»Θ§ΙΪ Ϋ»γœ¬ΘΚ
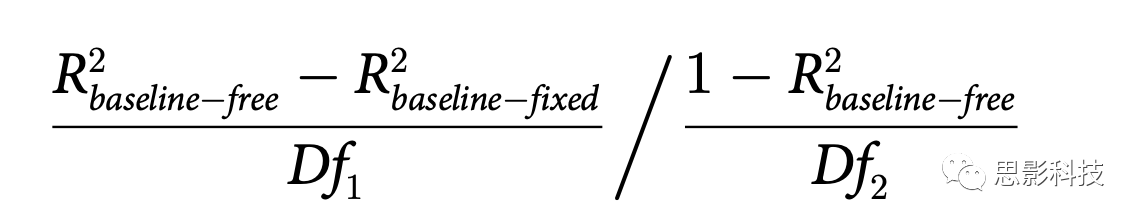
Τδ÷–R2baseline-free ΚΆR2baseline-fixedΖ÷±π¥”ΈόΜυœΏΡΘ–ΆΚΆΜυœΏΙΧΕ®ΡΘ–ΆΘ®Άξ’ϊΡΘ–ΆΚΆΦρΜ·ΡΘ–ΆΘ©ΒΡΡβΚœ÷–ΜώΒΟΓΘDf1 «Άξ’ϊΡΘ–Ά÷–ΒΡΉ‘”…≤Έ ΐ ΐ(24ΘΚ6 b's, 6 Gr's, 6 Gc's, ΚΆ 6 q'sΘ§Ζ÷±π”Ο”ΎΦ·÷–ΡΩ±ξΓΔΦ·÷–Ζ«ΡΩ±ξ“‘ΦΑ‘Ύ‘γΤΎΚΆΚσΤΎ―ΒΝΖΫΉΕΈΒΡΖ÷…ΔΉΔ“βΝΠΧθΦΰ)Φθ»ΞΦρΜ·ΡΘ–Ά÷–ΒΡΉ‘”…≤Έ ΐ ΐ(19: 6 Gr's, 6 Gc's,ΚΆ 6 q'sΖ÷±π «Ε‘”Ύ‘γΤΎΚΆΚσΤΎ―ΒΝΖΫΉΕΈΒΡΦ·÷–ΚΆΖ÷…Δ“‘ΦΑΡΩ±ξΚΆΖ«ΡΩ±ξΧθΦΰΘ§“‘ΦΑ‘ΎΥυ”–6Ηω Β―ιΧθΦΰ÷–Ι≤œμΒΡ1b)ΓΘDf2 «Ιέ≤λ¥Έ ΐ(36ΘΚ6Ε‘±»Ε»Υ°ΤΫ≥Υ“‘6Ηω Β―ιΧθΦΰ)Φθ»ΞΆξ’ϊΡΘ–Ά÷–ΒΡΉ‘”…≤Έ ΐ ΐΝΩ(24)Φθ1ΓΘFΖ÷≤Φ”Ο”ΎΙάΦΤΆξ’ϊΡΘ–Ά”κΦρΜ·ΡΘ–Ά≤ΜΆ§ΒΡœ‘÷χ–‘ΓΘΕ‘”ΎP1 ΐΨίΘ§ΈόΜυœΏΡΘ–Ά≤Δ≤ΜΟςœ‘”≈”ΎΜυœΏΙΧΕ®ΡΘ–ΆΘ§“ρ¥ΥΈ“Ο«÷ΜΤάΙάΝΥ Ι”ΟΙΧΕ®ΜυœΏ≤Έ ΐΒΡΖΫ≥Χ1ΙάΦΤΒΡΉνΦ―ΡβΚœ≤Έ ΐΒΡœ‘÷χ–‘ΓΘΝμ“ΜΖΫΟφΘ§Ε‘”ΎLPD ΐΨίΘ§ΈόΜυœΏΡΘ–ΆΟςœ‘ΗϋΚΟΘ§“ρ¥ΥΈ“Ο« Ι”Ο¥χ”–Ή‘”…”≈Μ·ΜυœΏ≤Έ ΐΒΡΙΪ Ϋ1ά¥±®ΗφΆ≥ΦΤΫαΙϊΓΘ
Ε‘”ΎΟΩ»’ΒΡP1ΚΆLPD ΐΨίΖ÷Έω(Φθ»ΞΜυœΏΚΆ≤ΜΦθ»ΞΜυœΏ)Θ§÷Ί≤…―υΚΆ÷ΊΡβΚœ≥Χ–ρΉώ―≠”κ…œ ωœύΆ§ΒΡΖΫΖ®ΓΘ
Ϋ®ΡΘΖΫΖ®
Έ“Ο«≤…”Οœ»«ΑΫ®ΝΔΒΡΜυ”ΎSDTΒΡΡΘ–Άά¥»ΖΕ®ΉΔ“β‘ω“φΚΆ‘κ“τ“÷÷ΤΒΡ≥ΧΕ»Θ§“‘Ϋβ Ά―ΒΝΖ‘γΤΎΚΆΚσΤΎ–ΡάμΈοάμΚΆ ¬ΦΰœύΙΊΒγΈΜ ΐΨί÷–ΒΡΉΔ“βΒς÷Τ÷°ΦδΒΡΙΊœΒΓΘΗΟΫ®ΡΘΩρΦήΜυ”Ύ’β―υΒΡΦΌ…ηΘ§Φ¥÷ΣΨθΝιΟτΕ»(d') ήΈΔΖ÷ΤΫΨυΖ¥”ΠΒΡœό÷ΤΘΚR(c+ΠΛc(c))-R(c)ΜρΠΛRΘ§”…2Ηω≤ΜΆ§ΒΡ¥ΧΦΛ(Φ¥±ξΉΦ¥ΧΦΛΚΆ≤β ‘¥ΧΦΛ)”’ΖΔΘ§≥ΐ“‘’β–©Ζ¥”ΠΒΡ÷πΗω ‘¥ΈΒΡ±δ“λ–‘(Π“)ΘΚ
d'= ΠΛR/Π“ (3)
Τδ÷–R « Ι”ΟNaka-RushtonΖΫ≥Χ(Β» Ϋ1)ΙάΦΤΒΡΦΌ…ηΒΡCRFΓΘΫαΚœd'ΚΆNaka-RushtonΖΫ≥ΧΘ§Ω…“‘ΗυΨίCRFΒΡΒΦ ΐ(Μρ–±¬ )ΙάΦΤΕ‘±»Ε»±φ±πψ–÷ΒΘ§»γœ¬ ΫΥυ ΨΘΚ
ΠΛc=ΠΛR/(dR/dc) (4)
Τδ÷–dR/dc «Μυ¥ΓCRFΒΡΒΦ ΐΓΘ
ΗυΨίΉΔ“β‘ω“φΡΘ–ΆΘ§‘Ύ…ώΨ≠‘Σ‘κ…υ(Π“)ΚψΕ®ΒΡΦΌ…ηœ¬(‘ω“φΡΘ–Ά)Θ§ΉΔ“β”’ΒΦΒΡΕ‘±»Ε»±φ±πψ–÷ΒΒΡΫΒΒΆΩ…“‘Άξ»Ϊ”ΟΜυ”Ύ ¬ΦΰœύΙΊΒγΈΜΒΡCRF(dR/dc)–±¬ ΒΡ‘ωΦ”ά¥Ϋβ ΆΓΘ»γΙϊCRF–±¬ ΒΡ‘ωΦ”ΝΩ≤ΜΉψ“‘Ϋβ Ά–ΡάμΈοάμTvCΘ®ψ–÷Β-Ε‘±»Ε»Θ§threshold versus contrastΘ©ΙΠΡήΒΡ±δΜ·Θ§‘ρ±Ί–κΦθ–ΓΠ“≤Έ ΐ“‘Ϋβ Ά–ΡάμΈοάμΕ‘±»ψ–÷ΒΒΡ±δΜ·Θ®‘κ…υΡΘ–ΆΘ©ΓΘ
Έ“Ο« Ι”Ο“‘œ¬≤Ϋ÷η”Π”ΟΉΔ“β‘ω“φΡΘ–ΆΘΚ Ήœ»Θ§Έ“Ο« Ι”ΟΨΏ”–Ήν–ΓΕΰ≥ΥΈσ≤νΙάΦΤΖΫΖ®ΒΡΕύœν ΫΚ· ΐ(Οί=3)ά¥ΙάΦΤ”Ο”Ύ’κΕ‘‘γΤΎΚΆΚσΤΎ―ΒΝΖΫΉΕΈΒΡΖ÷…ΔΉΔ“βΚΆΦ·÷–ΉΔ“βΒΡ–ΡάμΈοάμ―ßΒΡTvCΚ· ΐΓΘΫ”œ¬ά¥Θ§Έ“Ο«ΫαΚœNaka-RushtonΖΫ≥ΧΚΆd'ΖΫ≥ΧΘ§‘Ύ––ΈΣ±μœ÷Ήν≤νΒΡ‘γΤΎ―ΒΝΖΫΉΕΈΘ§ΡΘΡβΝΥΖ÷…ΔΉΔ“βΧθΦΰœ¬Μυ”ΎP1≤®ΒΡCRFΓΘ–η“ΣΉΔ“βΒΡ «Θ§‘Ύ÷ς“ΣΖ÷Έω÷–Θ§Ζ÷…ΔΉΔ“β ΐΨί «Μυ”ΎΖ÷…ΔΡΩ±ξΚΆΖ÷…ΔΖ«ΡΩ±ξ÷°ΦδΩγ‘ΫΝΫΗω¥ΧΦΛΦδΗτΒΡΤΫΨυ÷ΒΓΘ
ΡβΚœ≥Χ–ρΩΣ Φ”ΎΫΪΙάΦΤΒΡCRFΒΡΒΎ“ΜΒψ(c0=0%)…η÷ΟΈΣΜυœΏ≤Έ ΐ(b)Θ§»γœ¬Υυ ΨΘΚ
R(c0) = b (5)
»ΜΚσΕ®“εœ¬“ΜΗωΕ‘±»Ε»(C1)»γœ¬ΘΚ
c1=c0+ΠΛc0 (6)
Τδ÷–ΠΛc0 «Ε‘±»Ε»ΈΣ0% ±ΒΡΕ‘±»Ε»ψ–÷ΒΓΘ“ρ¥ΥΘ§ Ι”Οd'ΖΫ≥Χ(Β» Ϋ2)ΙάΦΤc1¥ΠΒΡœλ”Π»γœ¬ΘΚ
R(c1)=b+Π“ (7)
ΦΌ…ηd'=1Θ§œ¬“ΜΗωΕ‘±»Ε»ΒΡΕ®“ε»γœ¬ΘΚ
Ci=ci-1+ΠΛci-1 (8)
Τδ÷–iΒ±«ΑΒϋ¥ζΘ§«“i «¥σ”Ύ1ΒΡΓΘ»ΜΚσ‘Ύci¥ΠΒΡœλ”Π±ΜΙάΦΤ»γœ¬ΘΚ
R(c1)=R(ci-1)+Π“ (9)
ΉνΚσΝΫΗω≤Ϋ÷η(Β» Ϋ8ΚΆ9)“Μ÷±≥÷–χΒΫCRF±ΜΆξ»ΪΙάΦΤΈΣ÷ΙΓΘΆ®ΙΐΉν–ΓΜ·‘ΎΖ÷…ΔΡΩ±ξΧθΦΰœ¬Μυ”ΎP1Ζυ÷ΒΒΡCRFΙέ≤β÷ΒΚΆ‘Λ≤β÷Β÷°ΦδΒΡΉν–ΓΕΰ≥ΥΈσ≤νά¥”≈Μ·ΜυœΏΚΆ‘κ…υ≤Έ ΐ(bΚΆΠ“)ΓΘΈΣΝΥΦλ―ιΟΩΗω―ΒΝΖΫΉΕΈP1 CRFΒΡΉΔ“β‘ω“φ±δΜ· «ΖώΩ…“‘Ϋβ ΆTvCΙΠΡήΒΡ±δΜ·Θ§Έ“Ο« Ι”Ο…œ ωΫ®ΡΘ≥Χ–ρΙάΦΤΝΥ‘γΤΎΚΆΚσΤΎ―ΒΝΖΫΉΕΈΦ·÷–ΡΩ±ξΧθΦΰœ¬ΒΡP1 CRFΘ§ΒΪbΚΆΠ“≤Έ ΐ «ΙΧΕ®ΒΡΘ§’β–©≤Έ ΐ «Μυ”Ύ‘γΤΎ―ΒΝΖΫΉΕΈ¥”Ζ÷…ΔΉΔ“βΧθΦΰΜώΒΟΒΡ÷ΒΕχΙΧΕ®ΒΡΓΘΫ”œ¬ά¥Θ§Έ“Ο«≤β ‘ΝΥ‘Ύ‘γΤΎΚΆΚσΤΎ―ΒΝΖΫΉΕΈ‘ –μΦ·÷–ΡΩ±ξΧθΦΰœ¬ΒΡ‘κ…υ≤Έ ΐ±δΜ· «ΖώΡήΙΜœ‘÷χΧαΗΏΜυ”ΎSDT(–≈Κ≈Φλ≤βάμ¬έ)ΒΡΡΘ–ΆΒΡ‘Λ≤β–ßΙϊΓΘΈΣΝΥ Βœ÷’β“ΜΒψΘ§Έ“Ο«≤…”Ο»γ…œΥυ ωΖΫΖ®‘ΎΦ·÷–ΡΩ±ξΧθΦΰœ¬ΙάΦΤP1 CRF(Ε‘±»œλ”ΠΚ· ΐ)Θ§÷Μ «Έ“Ο«‘ –μΠ“≤Έ ΐΉ‘”…±δΜ·“‘’“ΒΫΉνΦ―ΡβΚœΓΘ»ΜΚσΘ§ΫΪ¥”ΚαΩγΖ÷…ΔΡΩ±ξΚΆΦ·÷–ΡΩ±ξΧθΦΰΙΧΕ®Π“≤Έ ΐΒΡ‘ω“φΡΘ–ΆΜώΒΟΒΡR2÷Β(ΦρΜ·ΡΘ–Ά)”κ Ι”ΟΫΒ‘κΡΘ–ΆΜώΒΟΒΡR2÷ΒΫχ––±»ΫœΘ§‘ΎΗΟΫΒ‘κΡΘ–Ά÷–“≤‘ –μΠ“≤Έ ΐ‘ΎΉΔ“βΧθΦΰ(Άξ»ΪΡΘ–Ά)÷°ΦδΉ‘”…±δΜ·ΓΘΗΟ±»Ϋœ Ι”Ο«ΕΧΉFΦλ―ιΘΚ
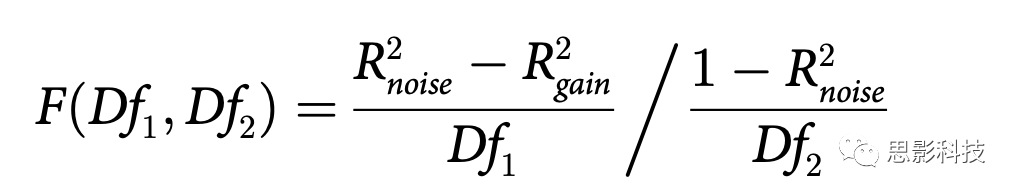
Τδ÷–R2gain ΚΆR2noise Ζ÷±π”…ΉΔ“β‘ω“φΚΆ‘κ…υΒς÷ΤΡΘ–Ά(ΦρΜ·ΡΘ–ΆΚΆΆξ»ΪΡΘ–Ά)ΡβΚœΒΟΒΫΓΘDf1 «Άξ’ϊΡΘ–Ά÷–ΒΡΉ‘”…≤Έ ΐΒΡ ΐΝΩ(3ΘΚ”Ο”ΎΦ·÷–ΉΔ“βΝΠΒΡΠ“Θ§”Ο”ΎΖ÷…ΔΉΔ“βΝΠΒΡΠ“Θ§“‘ΦΑ‘ΎΉΔ“βΝΠΧθΦΰ÷°ΦδΙ≤œμΒΡb)Φθ»ΞΦρΜ·ΡΘ–Ά÷–Ή‘”…≤Έ ΐΒΡ ΐΝΩ(2ΘΚ‘ΎΉΔ“βΝΠΧθΦΰ÷°ΦδΙ≤œμΒΡΠ“ΚΆb)ΓΘDf2 «Ιέ≤λ¥Έ ΐ(12ΘΚ6Ε‘±»Ε»Υ°ΤΫ≥Υ“‘2ΗωΉΔ“βΧθΦΰ)Φθ»ΞΆξ’ϊΡΘ–Ά÷–Ή‘”…≤Έ ΐΒΡ ΐΝΩ(3)Φθ1ΓΘFΖ÷≤Φ”Ο”ΎΙάΦΤΆξ’ϊΡΘ–Ά”κΦρΜ·ΡΘ–Ά≤ν“λœ‘÷χ–‘ΒΡΗ≈¬ ΓΘ
ΈΣΝΥ≤β ‘‘ω“φΡΘ–ΆΜρ‘κ…υΡΘ–Ά «ΖώΡήΗϋΚΟΒΊΫβ ΆΟΩΧλΒΡ––ΈΣ ΐΨίΚΆP1 ΐΨίΘ§Έ“Ο«≤…”ΟΝΥ»γ…œΥυ ωΒΡΜυ”ΎSDTΒΡΫ®ΡΘΩρΦήΓΘ
≥ΐΝΥΜυ”ΎSDTΒΡ‘ω“φΚΆ‘κ…υΡΘ–ΆΆβΘ§Έ“Ο«ΜΙ≤…”ΟΝΥ“Μ÷÷ΗΏ–ßΫβ¬κΡΘ–ΆΒΡ±δΧεΘ§Ω¥Ω¥Υϋ «ΖώΡήΗϋΚΟΒΊΫβ ΆP1≥…Ζ÷÷–ΒΡΉΔ“βΒς÷Τ”κΩγ―ΒΝΖΫΉΕΈΒΡ––ΈΣ ΐΨί÷°ΦδΒΡΝΣœΒΓΘΈΡ’¬÷ς“Σ≤…”Ο“Μ÷÷Μυ”ΎΉν¥σ≥ΊΙφ‘ρ(max-pooling,ΨΆ «‘Ύ“ΜΗωΙΧΕ®¥σ–ΓΒΡΜ§Ε·¥ΑΩΎ÷–»ΓΉν¥σ÷Β)ΒΡΗΏ–ßΫβ¬κΡΘ–ΆΒΡ±δΧεΘ§“‘Ϋβ ΆΚσΤΎ―ΒΝΖΫΉΕΈΒΡ––ΈΣΓΘ
ΫαΙϊ
––ΈΣΫαΙϊ
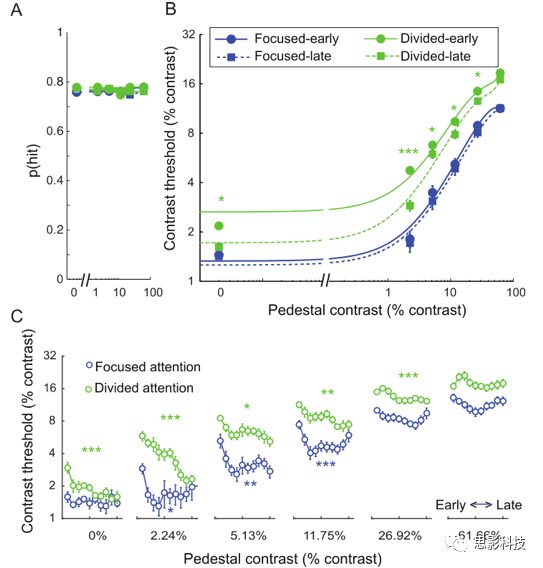
ΆΦ2BΥΒΟςΝΥΦ·÷–ΉΔ“βΝΠΚΆΖ÷…ΔΉΔ“βΝΠΧθΦΰœ¬“‘ψ–÷Β-Ε‘±»Ε»[threshold versus contrast Θ§TvC]«ζœΏΓΘ’βάοΗχ≥ωΒΡ––ΈΣ ΐΨί±ΜΖ÷≥…‘γΤΎΚΆΚσΤΎΝΫΗω―ΒΝΖΫΉΕΈΓΘΕ‘±»Ε»ψ–÷ΒΠΛcΥφΉ≈Ε‘±»Ε»ΒΡ‘ωΦ”Εχ‘ωΦ”ΘΜ”κΖ÷…ΔΉΔ“βΝΠœύ±»Θ§Φ·÷–ΉΔ“βΒΡΠΛcΦθ…ΌΓΘ’β–©ΉΔ“βΒςΫΎΖΔ…ζ‘ΎΥυ”–ΒΡΕ‘±»Ε»Υ°ΤΫΓΘ¥ΥΆβΘ§‘ΎΝΫΗω―ΒΝΖΫΉΕΈ÷–Θ§Ε‘ΠΛcΕΦ”–œ‘÷χΒΡΉΔ“β–ß”ΠΓΘΉνΚσΘ§―ΒΝΖ“≤ΫΒΒΆΝΥΠΛcΓΘ’β÷÷Ε‘ΠΛcΒΡ―ΒΝΖ–ßΙϊ÷ς“Σ «”…”Ύ‘Ύ0%-26.92%Ε‘±»Ε»Υ°ΤΫΒΡΖ÷…ΔΉΔ“β¥ΧΦΛœ¬ΓΘ
ΆΦ2 ––ΈΣΫαΙϊΓΘ
(A)‘ΎΥυ”–ΧθΦΰœ¬Θ§Οϋ÷–¬ ΕΦΙΧΕ®‘Ύ76%(d'=~1)ΓΘ
(B)‘ΎΦ·÷–ΚΆΖ÷…ΔΉΔ“βΧθΦΰœ¬Θ§Υυ”–ΜυΕ‘±»Ε»Υ°ΤΫ¥ΧΦΛΒΡ–ΡάμΈοάμΕ‘±»±φ±πψ–÷Β(ΠΛc)Ζ÷ΈΣ‘γΤΎΚΆΆμΤΎ―ΒΝΖΫΉΕΈΓΘ
(C)”κΖ÷…ΔΉΔ“βΧθΦΰœύ±»Θ§Φ·÷–ΉΔ“βΧθΦΰœ¬ΒΡ÷π»’ΠΛc ΐΨίœ‘ Ψ≥ωΗϋΩλΒΡ―ßœΑΥΌ¬ ΓΘ
ΈΣΝΥ‘ΎΗϋΨΪœΗΒΡ ±Φδ≥ΏΕ»…œΦλ―ιΉΔ“βΚΆ―ΒΝΖ–ßΙϊΘ§Έ“Ο«ΫΪ––ΈΣΒΡΕ‘±»±φ±πψ–÷Β ΐΨίΖ÷άύΒΫ10Ηω ±ΦδΕΈΓΘ≈δΕ‘tΦλ―ι±»ΫœΝΥΦ·÷–ΉΔ“βΚΆΖ÷…ΔΉΔ“βΧθΦΰœ¬Υυ”–Ε‘±»Υ°ΤΫΒΡ ΐΨίΘ§ΖΔœ÷Υυ”– ±ΦδΕΈΒΡ––ΈΣΕ‘±»ψ–÷ΒΨυ¥φ‘Ύœ‘÷χ≤ν“λΓΘ¥ΥΆβΘ§Έ“Ο«ΜΙΖΔœ÷Θ§Ζ÷…ΔΉΔ“βΚΆΦ·÷–ΉΔ“βΨυ¥φ‘Ύ―ßœΑ–ß”ΠΓΘΦ·÷–ΉΔ“βœ¬ΒΡ―ßœΑ–ß”ΠΒΡΖΔ…ζΥΌΕ»±»Ζ÷…ΔΉΔ“βœ¬“ΣΩλΒΟΕύΓΘΈΣΝΥΤάΙά’β–©―ßœΑ–ßΙϊΘ§Έ“Ο«Ε‘Ζ÷…ΔΉΔ“βΝΠΚΆΦ·÷–ΉΔ“βΝΠΧθΦΰœ¬Θ§“‘ΧλΉςΈΣΕ‘œσΡΎ“ρΥΊΕ‘ΟΩΗωΕ‘±»Υ°ΤΫΒΡ ΐΨίΫχ––ΝΥΒΞΕάΒΡΒΞœρ÷ΊΗ¥≤βΝΩΖΫ≤νΖ÷ΈωΓΘ‘ΎΖ÷…ΔΉΔ“βΧθΦΰœ¬Θ§ΜυΕ‘±»Ε»ΒΡ―ΒΝΖ–ßΙϊ‘Ύ0.014%~26.92%÷°Φδœ‘÷χΘ§Ε‘61.66%ΒΡΜυΕ‘±»Ε»ΟΜ”–œ‘÷χΒΡ―ΒΝΖ–ßΙϊΓΘ‘ΎΦ·÷–ΉΔ“βΒΡΧθΦΰœ¬Θ§Έ“Ο«ΖΔœ÷÷Μ”–ΜυΕ‘±»Ε»‘Ύ2.24%-11.75%÷°ΦδΒΡ―ΒΝΖ–ßΙϊœ‘÷χΓΘΦ·÷–ΉΔ“βΧθΦΰœ¬―ΒΝΖΕ‘––ΈΣΒΡ”Αœλ÷ς“ΣΖΔ…ζ‘ΎΒΎ“ΜΧλΚΆΒΎΕΰΧλ÷°ΦδΘ§“ρ¥Υ±»Ζ÷…ΔΉΔ“βΧθΦΰœ¬ΒΡ―ΒΝΖ–ß”Πά¥ΒΟΗϋ‘γΓΔΗϋΆΜ»ΜΓΘ
EEG ΫαΙϊΘΚ“ΜΑψΖ÷ΈωΖΫΖ®
ΈΣΝΥΦλ≤ι”…ΉΔ“βΚΆΖ«ΉΔ“β¥ΧΦΛ“ΐΤπΒΡΤΛ≤ψΖ¥”ΠΘ§Έ“Ο«Ή®ΉΔ”ΎΝΩΜ·ΒΎ“ΜΚΆΒΎΕΰ¥ΧΦΛ≥ œ÷ΦδΗτΒΡP1Ζ¥”ΠΒΡΤΫΨυ÷ΒΓΘ‘γΤΎ ”ΨθœΒΆ≥‘ΎΆβ≤Ω¥ΧΦΛΚΆΥϋΟ«ΒΡΤΛ≤ψ±μ Ψ÷°Φδ”–“ΜΗωΕ‘≤ύ”≥…δΘ§’β―υ‘ΎΉσ≤ύ ”“Α÷–≥ωœ÷ΒΡ¥ΧΦΛ‘Ύ”“≤ύ’μ“ΕΤΛ÷ ÷–ΜΫΤπΖ¥”ΠΘ§Ζ¥÷°“ύ»ΜΓΘΈΣΝΥΗϋΚΟΒΊΖ÷άκ”κΩ’ΦδΉσ≤ύΚΆ”“≤ύ≥ωœ÷ΒΡ¥ΧΦΛœύΙΊΒΡ¥ΧΦΛ”’ΖΔΖ¥”ΠΘ§Έ“Ο« Ήœ»‘Ύ Β―ι÷–Ε‘Ε‘≤ύ ”“Α÷–ΒΡΕ‘±»Ε»ΈΣ0%ΒΡΗ––Υ»ΛΒγΦΪΚΆΆ§≤ύ ”“Α÷–ΒΡΖ«ΝψΕ‘±»Ε»¥ΧΦΛΒΡERPsΫχ––ΝΥΤΫΨυΓΘΫ”œ¬ά¥Θ§Έ“Ο«¥”Ε‘≤ύ ”“Α≥ œ÷Ζ«ΝψΕ‘±»Ε» ”Ψθ¥ΧΦΛΒΡ ‘―ιΒΡ ¬ΦΰœύΙΊΒγΈΜ÷–Φθ»Ξ’βΗω ¬ΦΰœύΙΊΒγΈΜ(ΆΦ3)ΓΘ’β÷÷ΦθΖ®Ζ÷±π’κΕ‘ΟΩΗωΕ‘≤ύ¥ΧΦΛΕ‘±»Ε»Υ°ΤΫΓΔΟΩΗωΉΔ“βΧθΦΰΚΆΟΩΗω―ΒΝΖΫΉΕΈ÷¥––ΓΘΗΟΖΫΖ®≤ΜΫω”–÷ζ”Ύ¥”ΥΪ≤ύ¥ΧΦΛ’σΝ–÷–Ζ÷άκ”κΒΞΗω¥ΧΦΛœύΙΊΒΡ”’ΖΔΖ¥”ΠΘ§Εχ«“ΜΙ”–÷ζ”ΎΩΊ÷Τ”κΉΔ“βœΏΥς≥ œ÷œύΙΊΒΡ»ΈΚΈΩ’Φδ…œΖ«ΧΊΕ®ΒΡ‘ΛΤΎ–ß”ΠΓΘΫ”œ¬ά¥Θ§ΈΣΝΥΫχ“Μ≤Ϋ»Ζ±ΘΫαΙϊΡΘ Ϋ≤ΜΜα ήΒΫΦθΖ®ΒΡ»ΥΈΣ”ΑœλΘ§Έ“Ο«ΜΙΕ‘ΟΜ”–ΜυœΏΦθΖ®ΒΡ ΐΨίΫχ––ΝΥΖ÷ΈωΘ§Ιέ≤λΒΫΝΥœύΥΤΒΡΫαΙϊΡΘ ΫΘ§¥”Εχ―ι÷ΛΝΥΜυœΏΦθΖ®ΒΡœΏ–‘ΦΌ…ηΓΘ
ΆΦ3. ¬ΦΰœύΙΊΒγΈΜ(ERP)ΦθΖ®ΒΡ Ψάΐ
(A)ΉσΝ–(Ήœ…Ϊ)ΘΚ61.66%Ε‘±»Ε»ΒΡΖ÷…ΔΖ«ΡΩ±ξ¥ΧΦΛ Ψ“βΆΦΘ§œ‘ Ψ‘ΎΉσΑκ ”“Α(≥ΤΈΣΗ––Υ»Λ¥ΧΦΛ)Θ§≤Δ”κ“‘Υυ”–≤ΜΆ§Ε‘±»Ε»≥ œ÷ΒΡΡΩ±ξ¥ΧΦΛ≈δΕ‘ΓΘ”“άΗ(ΚΎ…Ϊ)ΘΚΉσΑκ ”“Α÷–0%Ε‘±»Ε»Ζ÷…ΔΖ«ΡΩ±ξ¥ΧΦΛ”κ”“Ακ ”“Α÷–ΒΡΆ§“ΜΉιΡΩ±ξ¥ΧΦΛ≈δΕ‘ΓΘ
(B)¥”Ε‘±»Ε»ΈΣ61.66%ΒΡΉσ≤ύΖ÷…ΔΖ«ΡΩ±ξ¥ΧΦΛ(AΘ§Ήσ≤ύΘΜBΘ§ΕΞ≤ΩΘ§–ιœΏΉœ…ΪΚέΦΘ)Υυ”’ΖΔΒΡERPΖ¥”Π÷–Φθ»ΞΉσ≤ύΕ‘±»Ε»ΈΣ0%ΒΡΖ÷…ΔΖ«ΡΩ±ξ¥ΧΦΛ(AΘ§”“≤ύΘΜBΘ§ΕΞ≤ΩΘ§ΚΎ…Ϊ–ιœΏ)Υυ”’ΖΔΒΡERPΖ¥”ΠΘ§ΒΟΒΫΦθ»ΞERPΖ¥”ΠΒΡΜυœΏ(BΘ§ΒΉ≤ΩΘ§ Β–ΡΉœ…ΪΙλΦΘ)ΓΘΥυ”–ΤδΥϊΕ‘±»÷–≥ œ÷ΒΡΗ––Υ»Λ¥ΧΦΛœύΙΊΒΡERPsΦΤΥψΨυΫχ––άύΥΤΒΡΦθΖ®ΓΘ
ΆΦ4. ¬ΦΰœύΙΊΒγΈΜ(ERP)ΫαΙϊΓΘ
(A) ‘γΤΎΚΆΆμΤΎ―ΒΝΖΫΉΕΈ”…Φ·÷–ΡΩ±ξΓΔΦ·÷–Ζ«ΡΩ±ξΓΔΖ÷…ΔΡΩ±ξΚΆΖ÷…ΔΖ«ΡΩ±ξΧθΦΰ“ΐΤπΒΡERPΘ§―’…ΪΒΡ“θ”Α¥ζ±μ¥ΧΦΛΒΡΕ‘±»Ε»Υ°ΤΫ(…ν…ΪΒΫΝΝ…Ϊ¥ζ±μΒΆΒΫΗΏΕ‘±»Ε»Υ°ΤΫ)ΓΘ
(B)P1≥…Ζ÷ΓΘ
(C)LPD≥…Ζ÷ΓΘ
(D)P1ΚΆLPDΒΡΒΊ–ΈΆΦ
―ΒΝΖΦθ»θ ”ΨθP1≥…Ζ÷ΒΡΉΔ“β‘ω“φ
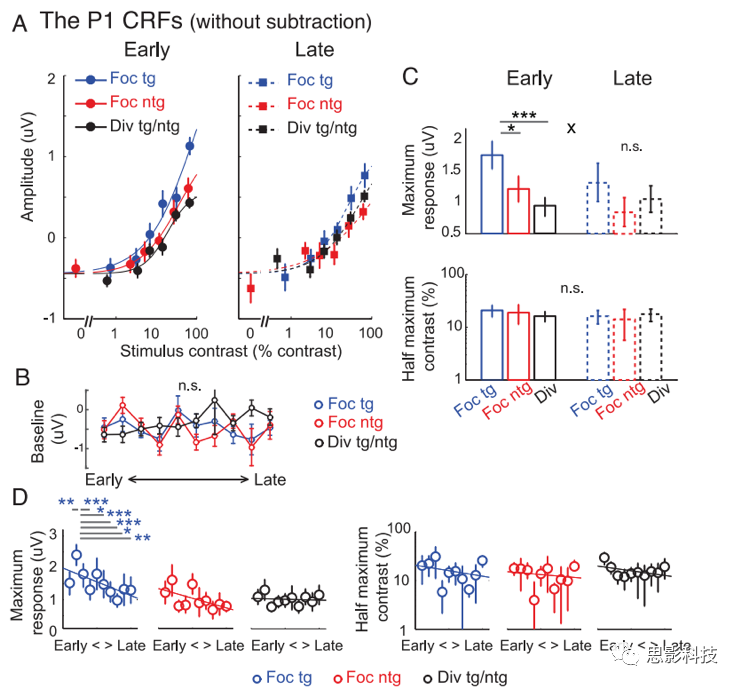
ΆΦ5A Ψ≥ωΝΥP1ΖυΕ»ΉςΈΣ¥ΧΦΛΕ‘±»Ε»ΒΡΚ· ΐΘ§ΈΣΦ·÷–ΡΩ±ξΓΔΦ·÷–Ζ«ΡΩ±ξ(ΜρΚω¬‘ΒΡ¥ΧΦΛ)ΚΆΖ÷…ΔΉΔ“βΧθΦΰ(‘ΎΖ÷…ΔΡΩ±ξΚΆΖ÷…ΔΖ«ΡΩ±ξΧθΦΰ…œΤΫΨυ)≤ζ…ζΕ‘±»œλ”ΠΚ· ΐ(CRF)ΓΘ»ΜΚσΘ§Έ“Ο« Ι”ΟNaka-RushtonΖΫ≥Χ(≤ΈΦϊΙΪ Ϋ1)ά¥±μ’ςΟΩ÷÷ΧθΦΰœ¬CRFΒΡ–ΈΉ¥Θ§“‘ΙάΦΤΉν¥σœλ”ΠΚΆΨωΕ®CRF―Ίx÷αΒΡΥ°ΤΫΈΜ÷ΟΒΡΑκΉν¥σΕ‘±»Ε»÷ΒΘ®ΆΦ5BΘ©ΓΘ

ΆΦ5.ΜυœΏΦθΖ®Ε‘P1≥…Ζ÷ΒΡΒς÷Τ
Θ®AΘ©P1Ε‘±»œλ”ΠΚ· ΐΓΘ
(B)ΈΣ(A)÷–Μυ”ΎP1ΒΡCRFΒΡœύ”ΠΉν¥σœλ”ΠΚΆΑκΉν¥σΕ‘±»Ε»ΓΘ
(C)‘Ύ«ΑΚσΝΫΗω―ΒΝΖΫΉΕΈΒΡΒΎ“ΜΚΆΒΎΕΰ¥ΧΦΛΤΎΦδΘ§”…Ζ÷…ΔΡΩ±ξΚΆΖ÷…ΔΖ«ΡΩ±ξ¥ΧΦΛΥυ”’ΖΔΒΡΜυ”ΎP1ΒΡCRFΓΘ
(D)ΆΦ(C)ΒΡœύ”ΠΉν¥σœλ”ΠΚΆΑκΉν¥σΕ‘±»Ε»ΓΘ
(E)”ΟΜυœΏΦθΖ®÷π»’Ζ÷ΈωP1 ΐΨίΓΘΉσΚΆ”“Ζ÷±π±μ ΨΙάΦΤΒΡΉν¥σœλ”ΠΚΆΑκΉν¥σΕ‘±»Ε»ΓΘ
Έ“Ο«‘Ύ―ΒΝΖ‘γΤΎΖΔœ÷Θ§Φ·÷–ΡΩ±ξΧθΦΰœ¬Μυ”ΎP1ΒΡCRFΒΡΉν¥σΖ¥”Πœ‘÷χ‘ωΦ” (ΆΦ5AΉσ≤ύΚΆ5BΕΞ≤Ω)ΓΘœύΖ¥Θ§‘ΎΚσΤΎ―ΒΝΖ÷–Θ§Έ“Ο«ΖΔœ÷Φ·÷–ΡΩ±ξΚΆΦ·÷–Ζ«ΡΩ±ξΧθΦΰ÷°ΦδΘ§Μρ’ΏΦ·÷–ΡΩ±ξΚΆΖ÷…ΔΉΔ“βΧθΦΰ÷°ΦδΟΜ”–≤ν“λ(ΆΦ5A”“ΚΆ5BΕΞ≤Ω)ΓΘ’β“ΜΉήΧεΡΘ Ϋ‘Ύ―ΒΝΖ(‘γΚΆΆμ)ΚΆΉΔ“βΧθΦΰ(Φ·÷–ΡΩ±ξΚΆΖ÷…ΔΉΔ“β)÷°Φδ≤ζ…ζΝΥœ‘÷χΒΡΫΜΜΞΉς”Ο(p=0.006)ΓΘ÷Ί“ΣΒΡ «Θ§Έ“Ο«ΖΔœ÷―ΒΝΖΚσΉΔ“β‘ω“φΒΡΦθ»θ «”…”Ύ”–―Γ‘ώΒΊΫΒΒΆΝΥΦ·÷–ΡΩ±ξΧθΦΰœ¬ΒΡΉν¥σP1Ζ¥”Π(p=0.013)ΓΘ―ΒΝΖœύΙΊΒΡP1≤®Ζυ±δΜ· «ΧΊΕ®”ΎΦ·÷–ΡΩ±ξΧθΦΰΒΡΘ§’β“Μ ¬ Β±μΟς―ΒΝΖ÷ς“Σ”Αœλ”κΦ·÷–ΉΔ“βΒςΕ»œύΙΊΒΡ…ώΨ≠ΒςΫΎΓΘ¥ΥΆβΘ§’β–©ΒςΫΎΒΡΧΊ“λ–‘±μΟςΘ§”κ―ΒΝΖœύΙΊΒΡP1ΖυΕ»ΒΡ±δΜ·≤Μ ή“ΜΑψΒΡΒΆΥ°ΤΫΗ–ΨθΜρ÷ΣΨθ―ßœΑ–ß”ΠΒΡ”ΑœλΘ§“ρΈΣ―ΒΝΖ≤ΜΜα”Αœλ”κ»ΈΚΈΤδΥϊΧθΦΰ(Φ¥Φ·÷–Ζ«ΡΩ±ξΧθΦΰΚΆΖ÷…ΔΉΔ“βΧθΦΰ)œύΙΊΒΡP1ΒΡ¥σ–ΓΓΘ
ΆΦ5eœ‘ ΨΝΥ÷π»’EEGΖ÷ΈωΒΡΫαΙϊΘ§ΗΟΫαΙϊ÷ß≥÷”κΖ÷ΈΣ‘γΤΎΚΆΚσΤΎ―ΒΝΖΫΉΕΈΒΡ≤πΖ÷ ΐΨίœύΆ§ΒΡΫα¬έΓΘ‘ΎΦ·÷–ΡΩ±ξΧθΦΰœ¬ΒΡ―ΒΝΖΒΡœ‘÷χ–ßΙϊ «”…ΒΎΕΰΧλCRFΉν¥σœλ”ΠΒΡœ‘÷χ‘ωΦ”ΆΤΕ·ΒΡΘ§Ά§“ΜΧλΘ§‘ΎΦ·÷–ΉΔ“βΧθΦΰœ¬ΒΡ––ΈΣ±μœ÷œ‘÷χΗΡ…Τ(ΆΦ2C)ΓΘ‘ΎΒΎ»ΐΧλΘ§Β±Φ·÷–ΉΔ“βΉ¥Χ§œ¬ΒΡ––ΈΣ±μœ÷ΩΣ ΦΈ»Ε® ±Θ§Ήν¥σP1Ζ¥”Πœύ”Πœ¬ΫΒΓΘ’β÷÷‘γΤΎ ”ΨθΖ¥”ΠΒΡ…œ…ΐΚΆœ¬ΫΒΡΘ Ϋ”κ÷ΣΨθ±μœ÷ΒΡΗΡ…ΤΚΆΈ»Ε®Ά§ ±ΖΔ…ζΓΘ―ΒΝΖΕ‘ΑκΉν¥σΕ‘±»Ε»ΟΜ”–±δΜ·Θ§ΫαΙϊ±μΟς―ΒΝΖ÷ς“Σ”ΑœλΉΔ“βΝΠΖ≈¥σ¥ΧΦΛ”’ΖΔΖ¥”ΠΒΡΉν¥σΖ¥”ΠΒΡ≥ΧΕ»ΓΘ
Έ“Ο«ΜΙΖ÷ΈωΝΥΟΜ”–Φθ»ΞΜυœΏΒΡΜυ”ΎP1ΒΡCRFΘ§ΫαΙϊ”κΦθ»ΞΜυœΏΜώΒΟΒΡΫαΙϊœύΥΤΓΘ¥”Εχ―ι÷ΛΝΥΜυœΏΦθΖ®ΒΡœΏ–‘ΦΌ…ηΓΘΈ“Ο« Ι”ΟΝΥ“ΜΗωΕνΆβΒΡΉ‘”…≤Έ ΐά¥Ϋβ ΆΧθΦΰ÷°ΦδΒΡΜυœΏ≤ν“λΓΘ’β“ΜΖ÷Έω±μΟςΘ§‘ –μΜυœΏ≤Έ ΐΉ‘”…±δΜ·≤ΔΟΜ”–œ‘÷χΧαΗΏΡβΚœ”≈Ε»Θ§Φ¥”κΜυ”ΎP1ΒΡCRFœύΙΊΒΡΜυœΏ≤Έ ΐ≤ΜΜαΥφΉ≈ΉΔ“βΝΠΜρ―ΒΝΖΕχΗΡ±δΓΘ¥ΥΆβΘ§”κΟΜ”–Φθ»ΞΜυœΏΒΡP1œύΙΊΒΡΑκΉν¥σΕ‘±»Ε»‘ΎΉΔ“βΧθΦΰΚΆ―ΒΝΖΫΉΕΈ÷–ΟΜ”–±δΜ·ΓΘ
ΆΦ6.ΟΜ”–Φθ»ΞΜυœΏΒΡP1≥…Ζ÷ΒΡΒς÷Τ
Ε®ΝΩΡΘ–Ά±μΟς―ΒΝΖΚ󥔑ω“φΒΫΫΒ‘κΒΡΉΣ±δ
ΉήΧεΕχ―‘Θ§P1ΒΡΫαΙϊ±μΟςΘ§ΉΔ“β‘ω“φ «‘Ύ―ΒΝΖ‘γΤΎ―Γ‘ώ–‘ΉΔ“βΒΡΜζ÷ΤΓΘ»ΜΕχΘ§―ΒΝΖΚσΤΎΉΔ“β‘ω“φΒΡ»± ß±μΟς―ΒΝΖΩ…ΡήΜαΗΡ±δ÷ß≥÷―Γ‘ώ–‘ΉΔ“βΒΡ…ώΨ≠Μζ÷ΤΓΘάΐ»γΘ§ΉνΫϋΒΡ―–ΨΩΧα≥ωΘ§Φ¥ Ι‘ΎΟΜ”–‘ω“φΒΡ«ιΩωœ¬Θ§ΫΒ‘κΜρ”––ßΒΡΫβ¬κΖΫΑΗ“≤Ω…“‘¥ΌΫχ―Γ‘ώ–‘Φ”ΙΛΓΘΈΣΝΥΤάΙά’β–©Χφ¥ζΒΡΫβ ΆΘ§Έ“Ο«≤…”ΟΝΥΜυ”ΎSDTΒΡΕ®ΝΩΫ®ΡΘΩρΦήά¥ΤάΙά‘ω“φΚΆ‘κ…υΒς÷ΤΕ‘––ΈΣΦ®–ßΒΡ”ΑœλΓΘΥφΚσΘ§Έ“Ο«ΜΙΤάΙάΝΥΗΏ–ßΫβ¬κΒΡΩ…ΡήΙ±œΉΓΘ
ΆΦ7.Μυ”Ύ–≈Κ≈Φλ≤βάμ¬έ(SDT)ΒΡΨΚ’υΉΔ“βΜζ÷Τ
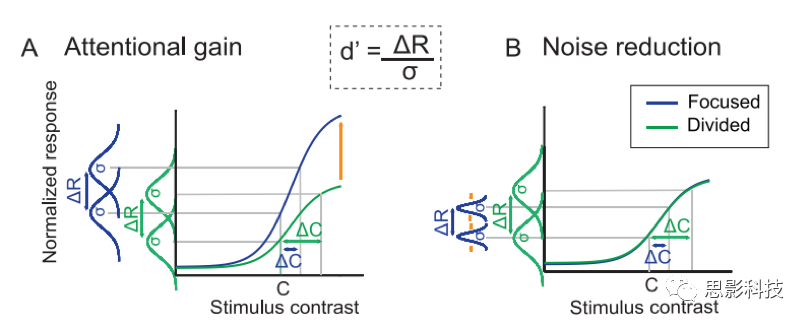
ΗυΨίSDTΘ§÷ΣΨθΟτΗ–Ε»(d')Ά®Ιΐ‘ωΦ”‘ω“φ(ΠΛR)ΚΆΫΒΒΆ‘κ…υ(Π“)ά¥‘ωΦ”ΓΘ
(A)ΉΔ“β‘ω“φΡΘ–ΆΦΌ…η―Γ‘ώ–‘ΉΔ“β‘ω«Ω‘γΤΎ ”ΨθΖ¥”ΠΒΡΖυΕ»(ΠΛR)Θ§¥”ΕχΒΦ÷¬÷ΣΨθΟτΗ–Ε»(d')ΒΡ‘ωΦ”ΓΘ
(B)ΫΒ‘κΡΘ–Ά»œΈΣΘ§ΉΔ“βΝΠΆ®ΙΐΫΒΒΆΠ“Φθ…ΌΝΥ÷πΗω ‘¥ΈΒΡ…ώΨ≠‘Σ±δ“λ–‘Θ§≤ΔΗΡ…ΤΝΥd'ΓΘΈ“Ο«ΦΌ…η―ΒΝΖΩ…ΡήΜαΕ®–‘ΒΊΗΡ±δ÷ß≥÷―Γ‘ώ–‘ΉΔ“βΒΡ…ώΨ≠Μζ÷ΤΘ§ΥφΉ≈ ±ΦδΒΡΆΤ“ΤΘ§¥”ΉΔ“β‘ωΦ”(A)ΉΣ±δΈΣ‘κ“τΫΒΒΆ(B)ΓΘ
»γΆΦ7ΚΆΙΪ Ϋ3Υυ ΨΘ§Μυ”ΎSDTΒΡΡΘ–ΆΦΌ…η÷ΣΨθΝιΟτΕ»(Μρd')”…ΝΫΗω≤ΜΆ§¥ΧΦΛ“ΐΤπΒΡΤΫΨυ…ώΨ≠Ζ¥”Π÷°ΦδΒΡ≤ν(ΠΛR)≥ΐ“‘’β–©Ζ¥”ΠΒΡ÷π¥Έ ‘¥Έ±δ“λ–‘(Π“)ά¥»ΖΕ®ΓΘΉΔ“β‘ω“φΩ…“‘Ά®Ιΐ‘ωΦ”ΠΛRά¥‘ωΦ”d'(ΆΦ7Θ§Ήσ)Θ§Εχ‘κ…υΫΒΒΆΩ…“‘Ά®ΙΐΫΒΒΆΠ“ά¥‘ωΦ”d'(ΆΦ7”“)ΓΘΈ“Ο« Ι”Ο’βΗωΡΘ–ΆΫΪΙέ≤λΒΫΒΡΕ‘±»Ε»±φ±πψ–÷Β”κΙέ≤λΒΫΒΡP1Ζ¥”ΠΒΡΒς÷Τ’ΐ ΫΝΣœΒΤπά¥ΓΘ
‘ω“φΡΘ–Ά‘Ύ‘γΤΎ―ΒΝΖΫΉΕΈ”––ßΒΊΫΪΕ‘±»Ε»ψ–÷ΒΒΡ±δΜ·ΚΆΜυ”ΎP1ΒΡCRFΒΡ–±¬ ΒΡ±δΜ·ΝΣœΒΤπά¥(±»ΫœΆΦ8AΉσ≤ύΒΡ«ύ…Ϊ«ζœΏΚΆάΕ…Ϊ‘≤»Π)Θ§Εχ‘κ…υΡΘ–Ά≤ΔΟΜ”–œ‘÷χΗΡ…Τ––ΈΣ ΐΨίΚΆΜυ”ΎP1ΒΡCRF÷°ΦδΒΡΡβΚœ(±»Ϋœ≥»…ΪΚΆ«ύ…Ϊ«ζœΏ)ΓΘ’β±μΟςΉΔ“β‘ω“φ≥δΖ÷Ϋβ ΆΝΥ‘γΤΎ―ΒΝΖ÷–…ώΨ≠ΚΆ––ΈΣ ΐΨί÷–ΉΔ“βΝΠΒςΫΎ÷°ΦδΒΡΙΊœΒΓΘ»ΜΕχΘ§‘ΎΚσά¥ΒΡ―ΒΝΖ÷–Θ§Φ¥ Ι‘ΎΦ·÷–ΉΔ“βΒΡ––ΈΣΖΫΟφ»‘»Μ”–ΗΡ…Τ(ΆΦ2B)Θ§P1ΖυΕ»ΒΡΉΔ“βΒςΫΎ(ΆΦ8AΒΡ”“≤ύ)“≤ΟΜ”–≥ωœ÷ΓΘ“ρ¥ΥΘ§‘ΎΚσΤΎ―ΒΝΖΫΉΕΈΘ§‘ω“φΡΘ–ΆΗΏΙάΝΥΜυ”ΎP1ΒΡCRF‘ΎΦ·÷–ΡΩ±ξΧθΦΰœ¬ΒΡ–±¬ (±»ΫœΆΦ8A”“≤ύ÷–ΒΡ«ύ…Ϊ«ζœΏΚΆάΕ…Ϊ‘≤»Π)ΓΘœύΖ¥Θ§”κ‘ω“φΡΘ–Άœύ±»Θ§‘Ύ≤ΜΆ§ΧθΦΰœ¬Θ§‘κ…υΡΘ–ΆΨυΧαΙ©ΝΥΗϋΚΟΒΡΡβΚœ(±»Ϋœ≥»…ΪΚΆ«ύ…Ϊ«ζœΏΘ§ΆΦ8A-8L)ΓΘ
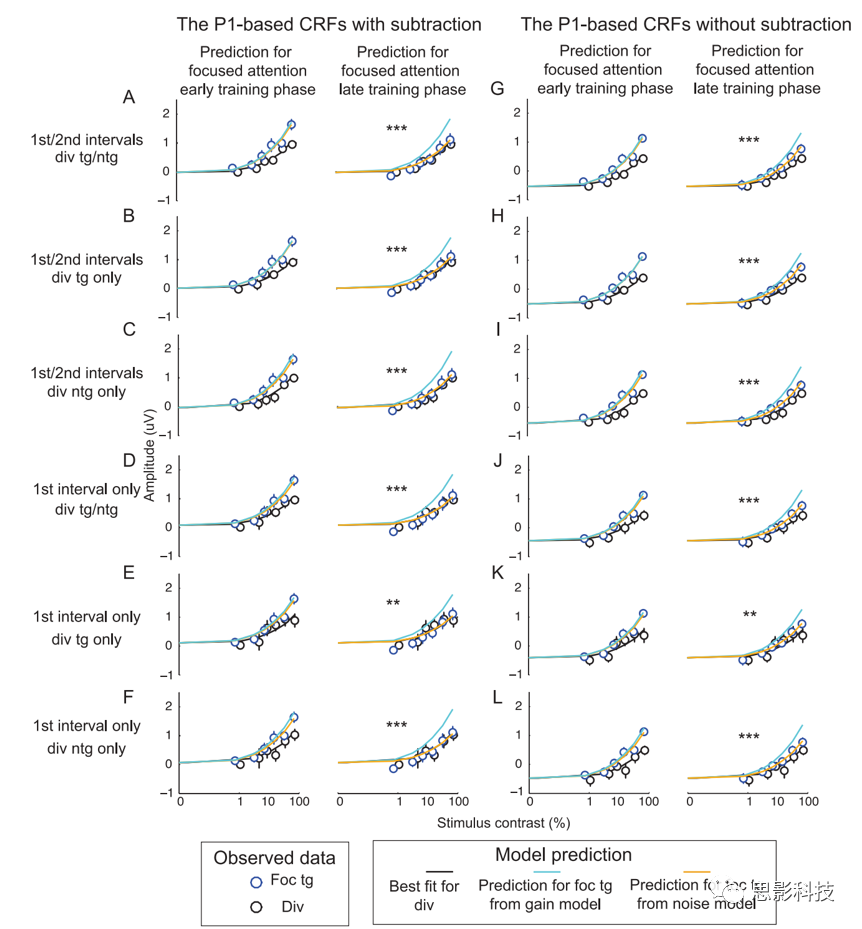
ΆΦ8. Ι‘ω“φΚΆ‘κ…υΡΘ–ΆΫΪ–ΡάμΈοάμ ΐΨίΚΆP1 ΐΨί±δΜ·ΒΡΝΣœΒΓΘ
ΗυΨίΖ÷…ΔΡΩ±ξΚΆΖ÷…ΔΖ«ΡΩ±ξΧθΦΰ(AΓΔDΓΔGΚΆJ)ΒΡΤΫΨυ÷ΒΘ§ΫωΗυΨίΖ÷…ΔΡΩ±ξΧθΦΰ(BΓΔEΓΔHΚΆK)Θ§ΫωΗυΨίΖ÷…ΔΖ«ΡΩ±ξΧθΦΰ(CΓΔFΓΔIΚΆL)Θ§¥”ΝΫΗω¥ΧΦΛΦδΗτ(A-CΚΆG-I)÷°ΦδΒΡΤΫΨυ÷ΒΘ§“‘ΦΑΫω¥”ΒΎ“ΜΗω¥ΧΦΛΦδΗτ(D-FΚΆJ-L)Θ§ΜώΒΟΖ÷…ΔΉΔ“βΧθΦΰœ¬Μυ”ΎP1ΒΡΕ‘±»Ζ¥”ΠΚ· ΐ(CRF)ΓΘ(A-F)œ‘ ΨΦθ»ΞΜυœΏΒΡ ΐΨίΘ§(G-L)œ‘ ΨΟΜ”–Φθ»ΞΜυœΏΒΡ ΐΨίΓΘ
Ι”Ο…œ ωΜυ”ΎSDTΒΡΫ®ΡΘΩρΦή÷π»’Ζ÷Έω––ΈΣΚΆP1 ΐΨίΩ…“‘ΖΔœ÷Θ§¥Ω‘ω“φΡΘ–ΆΩ…“‘Ϋβ Ά«Α2Χλ÷–P1œλ”ΠΚΆ––ΈΣ÷°ΦδΒΡΝΣœΒΘ§Εχ–η“ΣΫΒ‘κΡΘ–Άά¥ΗϋΚΟΒΊΫβ Ά¥σ≤ΩΖ÷Κσ–χΧλ ΐ÷–P1ΚΆ––ΈΣ ΐΨί÷°ΦδΒΡΝΣœΒΓΘΝμΆβΘ§Φθ»ΞΚΆ≤ΜΦθ»ΞΜυœΏΒΡP1 ΐΨίΒΡΫ®ΡΘ ΐΨί‘Ύ÷ ΝΩ…œ «œύΥΤΒΡΓΘ
―ΒΝΖ≤ΜΜαœς»θΆμΤΎ’ΐΤΪΉΣ(LPD)≥…Ζ÷ΒΡΉΔ“β‘ω“φ
’βΗωΫœΆμΒΡERP≥…Ζ÷LPD±Μ»œΈΣ «Η–ΨθΚσΦ”ΙΛ÷Η±ξΘ§άΐ»γΨω≤ΏΙΐ≥Χ÷–ΒΡ÷ΛΨίΤάΙάΓΘ»γΆΦ10AΥυ ΨΘ§Έ“Ο«ΖΔœ÷LPD‘Ύ‘γΤΎΚΆΚσΤΎΒΡ―ΒΝΖΫΉΕΈΕΦ”–Κή«ΩΒΡΉΔ“β‘ω“φΓΘΜυ”ΎLPDΒΡCRFΒΡΉν¥σœλ”ΠΘ§Φ·÷–ΡΩ±ξΧθΦΰΉνΗΏΘΜΑκΉν¥σΕ‘±»Ε»Θ§Φ·÷–Ζ«ΡΩ±ξΧθΦΰΉνΗΏΘ§‘γΤΎΚΆΚσΤΎ―ΒΝΖΫΉΕΈΨυΈό±δΜ·ΓΘΕ‘LPD ΐΨίΒΡ÷π»’Ζ÷Έωœ‘ ΨΘ§‘Ύ’ϊΗω―ΒΝΖΙΐ≥Χ÷–Θ§P1-CRFΒΡΉν¥σΖ¥”ΠΚΆΑκΉν¥σΕ‘±»Ε»ΒΡΉΔ“βΒςΫΎ“≤ «œύΕ‘Έ»Ε®ΒΡΘ®ΆΦ10EΘ©ΓΘ
‘Ύ―ΒΝΖΚσΤΎΘ§Έ“Ο«»Ζ ΒΙέ≤λΒΫΝΥ‘ΎΒΎΕΰ¥ΧΦΛ≥ωœ÷ΒΡΖ÷…ΔΡΩ±ξΚΆΖ÷…ΔΖ«ΡΩ±ξΧθΦΰœ¬ΒΡLPDΖ¥”ΠΒΡ≤ν“λΓΘ¥ΥΆβΘ§Έ“Ο«ΜΙΖΔœ÷¥ΧΦΛάύ–Ά(Ζ÷…ΔΡΩ±ξΚΆΖ«ΡΩ±ξ)ΓΔ¥ΧΦΛΦδΗτ(ΒΎ“ΜΚΆΒΎΕΰ)ΚΆ―ΒΝΖ(‘γΚΆΆμ)‘ΎΜυ”ΎLPDΒΡCRFΒΡΑκΉν¥σΕ‘±»Ε»…œ¥φ‘Ύœ‘÷χΒΡ3œρΫΜΜΞΉς”Ο(p=0.001Θ§)ΓΘ÷π»’Ρ‘ΒγΖ÷Έω”κ¥ΥΖΔœ÷“Μ÷¬ΓΘΖ÷…ΔΡΩ±ξΚΆΖ÷…ΔΖ«ΡΩ±ξΒΡLPDΖ¥”Π(ΆΦ10CΓΔ10DΚΆ10F)“≤±μΟςΘ§÷ΣΨθΚσΙΐ≥ΧΩ…“‘Ά®Ιΐ―ΒΝΖΗΡ…ΤΘ§Εχ≤ΜΜαΑιΥφ‘γΤΎΗ–ΨθΖ¥”ΠΒΡ±δΜ·(ΆΦ5CΚΆ5D)ΓΘ
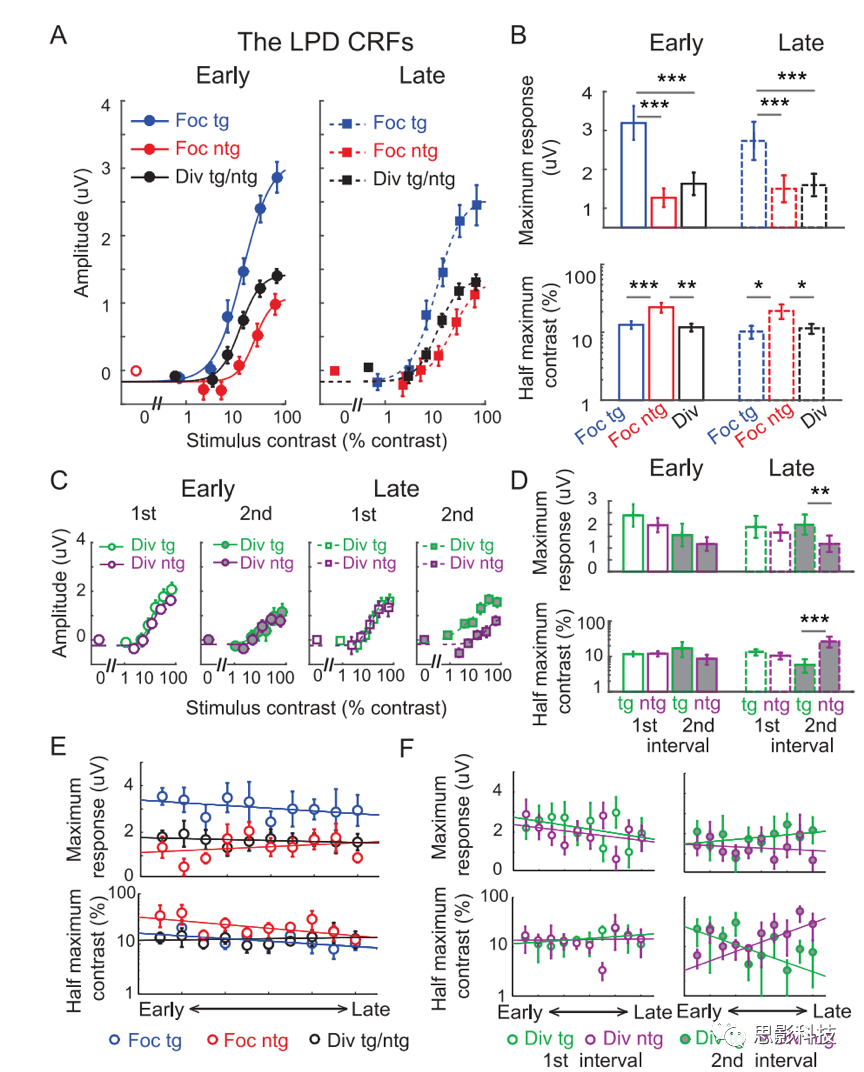
ΆΦ10.ΜυœΏΦθΖ®Ε‘ΆμΤΎ’ΐΤΪΉΣ(LPD)≥…Ζ÷ΒΡΒς÷Τ
(A)LPDΕ‘±»œλ”ΠΚ· ΐ(CRF)ΓΘ
(B)œύ”ΠΒΡΉν¥σœλ”ΠΚΆ(A)Υυ ΨCRFΒΡΑκΉν¥σΕ‘±»Ε»≤Έ ΐΓΘ
(C)‘Ύ‘γΤΎΚΆΚσΤΎ―ΒΝΖΫΉΕΈΒΡΒΎ“ΜΚΆΒΎΕΰ¥ΧΦΛΤΎΦδΘ§”…Ζ÷…ΔΡΩ±ξΚΆΖ«ΡΩ±ξ¥ΧΦΛΥυ”’ΖΔΒΡΜυ”ΎLPDΒΡCRFΓΘ
(D)ΆΦ(C)Υυ ΨCRFΒΡœύ”ΠΉν¥σœλ”ΠΚΆΑκΉν¥σΕ‘±»Ε»≤Έ ΐΓΘ
(E)Φθ»ΞΜυœΏΒΡLPD ΐΨίΒΡ÷π»’Ζ÷ΈωΘ§ΕΞ≤ΩΚΆΒΉ≤Ω±μ ΨΙάΦΤΒΡΉν¥σœλ”ΠΚΆΑκΉν¥σΕ‘±»Ε»≤Έ ΐΓΘ
(F)ΈΣ(C)ΚΆ(D)Υυ ΨΒΡLPD ΐΨίΒΡ÷π»’Ζ÷ΈωΓΘ
ΟΜ”–Φθ»ΞΜυœΏΒΡLPD(’ΐΤΪΉΣ) ΐΨίΘΚ«ΕΧΉΡΘ–Ά±»ΫœΖ÷Έω÷Λ ΒΘ§”κΙΧΕ®ΜυœΏ≤Έ ΐœύ±»Θ§‘ –μΜυœΏ≤Έ ΐΉ‘”…±δΜ·œ‘÷χΧαΗΏΝΥΡβΚœ”≈Ε»ΓΘ’β”κP1≥…Ζ÷ «≤ΜΆ§ΒΡΓΘ‘ΎLPD≥…Ζ÷÷–Θ§ΜυœΏΜνΕ·ΒΡ―ΒΝΖ–ßΙϊ±μΟςΘ§Ζ÷…ΔΉΔ“βΧθΦΰœ¬ΒΡΗΡ…Τ––ΈΣΖΔ…ζ‘Ύ÷ΣΨθΚσΫΉΕΈΘ§Εχ≤Μ «‘ΎΗ–ΨθΜρ÷ΣΨθΒΡ‘γΤΎΫΉΕΈΓΘ
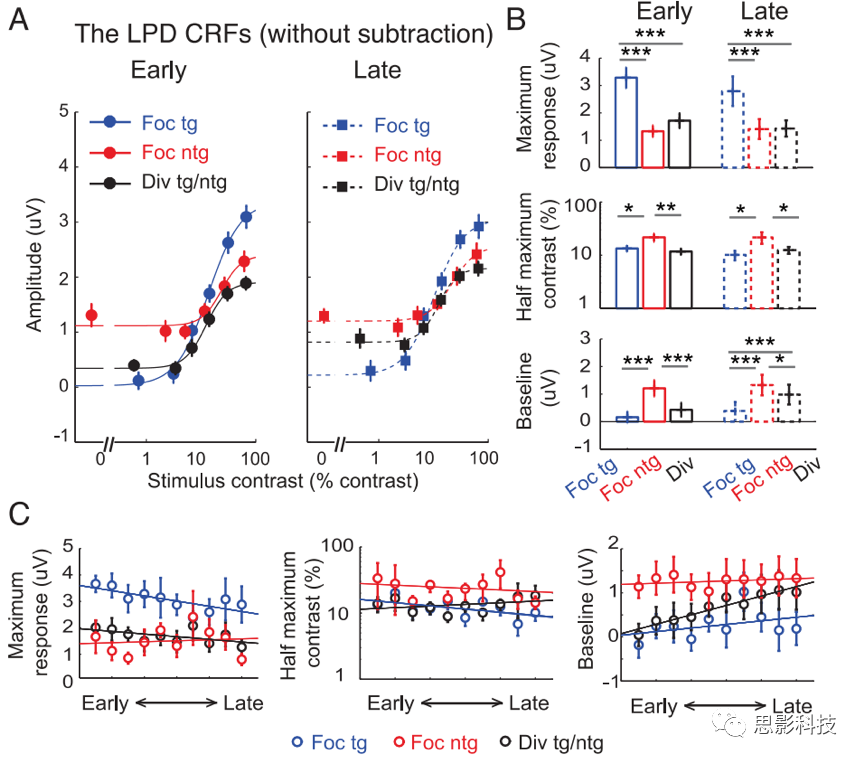
ΆΦ11.ΟΜ”–Φθ»ΞΜυœΏΕ‘ΆμΤΎ’ΐΤΪΉΣ(LPD)≥…Ζ÷ΒΡΒς÷Τ
Ε‘”Ύ±ΨΈΡΈ“Ο«Ω…“‘”–»γœ¬ΥΦΩΦΘΚ
±ΨΤΣΈΡ’¬Ϋω≤…”ΟΝΥ“ΜΗωΫœΈΣΨ≠ΒδΒΡ―Γ‘ώ–‘ΉΔ“βERPΖΕ ΫΘ§ΡήΙΜΖΔ±μ‘Ύ“Μ«χ‘”÷ΨPLOS BIOLOGY…œΘ§±μΟςΝΥΉς’ΏΡήΙΜΆ®Ιΐ»ΪΟφΕχ…ν»κΒΡΖ÷ΈωΘ§≤ϊ ω«ε≥ΰ―ΒΝΖΒΡ≥÷–χ ±Φδ»γΚΈΫΪ ”ΨθΤΛ≤ψ”κ––ΈΣΒΡΉΔ“βΒςΫΎΝΣœΒΤπά¥Θ§»γΚΈ¥”ΉΔ“β‘ω“φΒΫ‘κ“τΫΒΒΆΒΡΜζ÷ΤΉΣ±δΘ§Έ“Ο«‘Ό“ΜΤπ αάμ“Μœ¬ΕΦ≤…”ΟΝΥΡΡ–©÷ς“ΣΒΡΖ÷Έω÷Η±ξΘΚ
Θ®aΘ©––ΈΣΖ÷ΈωΘΚ
Θ®1Θ©ΫαΚœΉΔ“βΧθΦΰ(Φ·÷–ΉΔ“βΚΆΖ÷…ΔΉΔ“β)ΓΔ―ΒΝΖΫΉΕΈ(‘γΤΎΚΆΚσΤΎ)ΚΆ¥ΧΦΛΕ‘±»(0%~61.66%)ΒΡΕ‘±»±φ±πψ–÷ΒΘ§Φλ―ι’β–©“ρΥΊΦΑΤδΫΜΜΞΉς”ΟΕ‘Ε‘±»±φ±πψ–÷ΒΒΡ÷ς–ß”ΠΓΘ
Θ®2Θ©÷π»’––ΈΣΖ÷ΈωΩΦ≤λΦ·÷–ΉΔ“βΚΆΖ÷…ΔΉΔ“βΧθΦΰœ¬Θ§―ΒΝΖΕ‘≤ΜΆ§Ε‘±»Ε»Υ°ΤΫ¥ΧΦΛΒΡΕ‘±»Ε»±φ±πψ–÷ΒΒΡ”ΑœλΓΘ
Θ®bΘ©EEGΖ÷ΈωΘΚ
Θ®1Θ©Η––Υ»ΛΒΡ¥ΧΦΛ“ΐΤπΒΡERPsΒΡΧα»ΓΘ§Φθ»ΞΜυœΏΒΡ ΐΨίVS≤ΜΦθ»ΞΜυœΏΒΡ ΐΨίΓΘ
Θ®2Θ©ΟΩΗωΉΔ“βΧθΦΰΓΔΟΩΗω¥ΧΦΛάύ–ΆΚΆΟΩΗω―ΒΝΖΫΉΕΈΧαΙ©Μυ”ΎP1ΒΡCRFΘ§“‘ΦΑΙάΦΤΉν¥σœλ”ΠΓΔΑκΉν¥σΕ‘±»Ε»ΓΘ
Θ®3Θ©”κP1œύΆ§ΧθΦΰΒΡΜυ”ΎLPDΒΡCRFΘ§“‘ΦΑΙάΦΤΉν¥σœλ”ΠΓΔΑκΉν¥σΕ‘±»Ε»ΓΘ
Θ®4Θ©‘γΤΎΚΆΆμΤΎΝΫΗωΫΉΕΈVS÷π»’ °ΗωΫΉΕΈΓΘ
Θ®5Θ©ΈόΜυœΏΡΘ–ΆVSΜυœΏΙΧΕ®ΡΘ–ΆΓΘ
Θ®cΘ©Μυ”ΎSDTΒΡ‘ω“φΚΆ‘κ…υΡΘ–ΆΘΚΫβ Ά―ΒΝΖ‘γΤΎΚΆΚσΤΎ–ΡάμΈοάμΚΆ ¬ΦΰœύΙΊΒγΈΜ ΐΨί÷–ΒΡΉΔ“βΒς÷Τ÷°ΦδΒΡΙΊœΒΓΘ
Θ®dΘ©ΗΏ–ßΫβ¬κΡΘ–ΆΒΡ±δΧεΘΚΜυ”ΎΉν¥σ≥ΊΙφ‘ρ(max-pooling,ΨΆ «‘Ύ“ΜΗωΙΧΕ®¥σ–ΓΒΡΜ§Ε·¥ΑΩΎ÷–»ΓΉν¥σ÷Β)ΒΡΗΏ–ßΫβ¬κΡΘ–ΆΒΡ±δΧεΘ§“‘Ϋβ ΆΚσΤΎ―ΒΝΖΫΉΕΈΒΡ––ΈΣΓΘ
Ά®ΙΐœΒΆ≥–‘Ζ÷ΈωΩ…“‘Ω¥≥ωΘΚ
Θ®1Θ©‘ΎΫΪ ΐΨίΖ÷ΈΣ‘γΤΎΚΆΚσΤΎΝΫ≤ΩΖ÷ΒΡΖ÷Έω÷–Θ§‘ΎΦ·÷–ΉΔ“βΧθΦΰœ¬Θ§―ΒΝΖΕ‘––ΈΣ±μœ÷ΒΡ”ΑœλΉν–ΓΓΘ»ΜΕχΘ§Β±Έ“Ο«÷π»’Φλ≤ι––ΈΣ ΐΨί ±ΖΔœ÷Θ§‘ΎΦ·÷–ΉΔ“βΝΠΒΡ«ιΩωœ¬Θ§––ΈΣ±μœ÷ΒΡ±δΜ·±»Ζ÷…ΔΉΔ“βΝΠΒΡ«ιΩωœ¬ΖΔ…ζΒΡΥΌΕ»“ΣΩλΒΟΕύΓΘΜυ”ΎP1ΒΡCRF‘ΎΦ·÷–ΡΩ±ξΧθΦΰœ¬ΒΡΉν¥σœλ”Π¥”ΒΎ“ΜΧλΒΫΒΎΕΰΧλ‘ωΦ”ΓΘP1Ζ¥”ΠΒΡ‘ωΦ”“≤”Γ÷ΛΝΥ––ΈΣ±μœ÷ΒΡœ‘÷χΗΡ…ΤΓΘ»ΜΕχΘ§P1Ήν¥σœλ”Π‘ΎΒΎΕΰΧλΚσΟςœ‘œ¬ΫΒΘ§––ΈΣ±μœ÷ΩΣ ΦΈ»Ε®ΓΘ
Θ®2Θ© ”ΤΛ≤ψ÷–…ώΨ≠‘ω“φΚΆ…ώΨ≠‘κ…υΒΡΉΔ“βΝΠœύΙΊ±δΜ·“άάΒ”Ύ≤ΜΆ§ΒΡ ήΧεΆ®¬ΖΓΘΉΔ“βΝΠ‘ω“φΖ≈¥σ «”…MΒ®Φν–Ά““θΘΒ®Φν ήΧεΒςΫΎΘ§Εχ”κΉΔ“βΝΠœύΙΊΒΡ‘κ“τΫΒΒΆ‘ρ «”…N-ΦΉΜυ-D-ΧλΕ§Α±Υα(NMDA) ήΧεΆ®¬ΖΒςΫΎΘ§’β «“ΜΧθΙΊΦϋΒΡ ήΧεΆ®¬ΖΘ§≤Έ”κ–μΕύ―ßœΑΚΆΦ«“δΙΐ≥ΧΘ§Αϋά®≥Λ ±≥Χ‘ω«ΩΓΘ
Θ®3Θ© Β ©ΝΥΜυœΏΦθΖ®Θ§¥”ΥΪ≤ύ¥ΧΦΛ÷–Ζ÷άκ≥ω”κΒΞΗω¥ΧΦΛœύΙΊΒΡ”’ΖΔΖ¥”ΠΘ§≤ΔΉν–ΓΜ·”…”ΎΉΔ“βœΏΥςΕχ≤ζ…ζΒΡΩ’ΦδΖ«ΧΊ“λ–‘‘ΛΤΎ–ß”ΠΓΘ’β÷÷ΦθΖ®ΦΌ…ηΡ‘ΒγΖ¥”Π «œΏ–‘ΉιΚœΒΡΓΘ¥ΥΆβΘ§Έ“Ο«Ιέ≤λΒΫ”–ΚΆΟΜ”–ΜυœΏΦθΖ®ΒΡΫαΙϊ‘Ύ÷ ΝΩ…œœύΥΤΒΡΡΘ ΫΘ§ΧΊ±π «‘ΎP1≥…Ζ÷ΒΡ«ιΩωœ¬ΓΘ
Θ®4Θ©”κP1ΫαΙϊœύΖ¥Θ§‘Ύ’ϊΗω―ΒΝΖΙΐ≥Χ÷–Ιέ≤λΒΫΝΥ“Μ÷¬ΒΡLPDΉΔ“β‘ω“φΒς÷ΤΓΘLPD”κΗ–ΨθΚσΨω≤ΏœύΙΊΒΡΦ”ΙΛΟή«–œύΙΊΘ§≤Δ”κ÷ν»γΖ¥”Π–≈–ΡΓΔ»ΈΈώΡ―Ε»ΚΆΨω≤Ώ ±ΦδΒ»“ρΥΊœύΙΊΓΘ’β–©ΖΔœ÷±μΟςΘ§Υδ»Μ‘γΤΎΗ–Ψθ–≈Κ≈ΒΡΉΔ“βΒς÷ΤΩ…“‘ΥφΉ≈―ΒΝΖΖΔ…ζ Β÷ –‘ΒΡ±δΜ·Θ§ΒΪΗ–÷ΣΚσΙΐ≥ΧΒΡ‘ω“φΒς÷ΤΥφΉ≈ ±ΦδΒΡΆΤ“Τ±Θ≥÷œύΕ‘Έ»Ε®ΓΘ
Θ®5Θ©‘Ύ≥ΛΤΎ―ΒΝΖΧθΦΰœ¬Θ§ ”Ψθ”’ΖΔP1≥…Ζ÷¥”ΉΔ“β‘ω“φΒΫΫΒΒΆ‘κ…υΘ§LPD≥…Ζ÷ΒΡΉΔ“β‘ω“φ≥÷–χ¥φ‘ΎΓΘ’β «”κ¥σΡ‘ΤΛ≤ψ≤ψΦΕΦδ–≈œΔ¥Ϊ δ–߬ ΒΡΉΔ“βΝΠœύΙΊΗΡ…Τ «“Μ÷¬ΒΡΘ§ ΙΒΟ‘γΤΎΫΉΕΈ‘Ύ―ΒΝΖΚσœρœ¬”Έ«χ”ρΧαΙ©ΗϋΈ»Ε®ΒΡ–≈œΔΓΘΉν÷Ί“ΣΒΡ «Θ§―ΒΝΖ≤ΜΜαΦρΒΞΒΊΗΡ±δΗ–Ψθ–≈Κ≈ΒΡ¥σ–ΓΜρΗ–Ψθ–≈Κ≈ΒΡΉΔ“βΝΠΒςΫΎΒΡ¥σ–ΓΘ§ΒΪ «Θ§―ΒΝΖΩ…“‘Ε®–‘ΒΊΗΡ±δ…ώΨ≠Ζ¥”ΠΚΆ––ΈΣΒΡΉΔ“βΒςΫΎ÷°ΦδΒΡΙΊœΒΓΘ
«κΈΔ–≈Υ―ΥςΓΑΥΦ”ΑΩΤΦΦΓ±ΙΊΉΔΈ“Ο«Θ§ΙΊΉΔΈ“Ο«ΒΡΙΪ÷ΎΚ≈Θ§ΜώΒΟΗϋΕύΨΪ≤ Ρ‘ΩΤ―ßΡΎ»ίΘ§»γ–η‘≠ΈΡΜρΕ‘ΥΦ”ΑΩΤΦΦΩΈ≥ΧΗ––Υ»Λ«κΧμΦ”ΈΔ–≈Κ≈siyingyxfΜρ19962074063

ΜΕ”≠δ·άάΥΦ”ΑΒΡ ΐΨί¥ΠάμΩΈ≥Χ“‘ΦΑ ΐΨί¥Πάμ“ΒΈώΫι…ήΓΘΘ®«κ÷±Ϋ”ΒψΜςœ¬ΈΡΈΡΉ÷Φ¥Ω…δ·άάΥΦ”ΑΩΤΦΦΤδΥϊΩΈ≥ΧΘ§ΜΕ”≠ΧμΦ”ΈΔ–≈Κ≈siyingyxfΜρ19962074063Ϋχ––Ή…―·Θ§»‘Ϋ” ή±®ΟϊΘ§ ή“Ώ«ι”Αœλ≤ΩΖ÷ΩΈ≥Χ ±ΦδΜρ”–Βς’ϊΘ§±®ΟϊΚσΈ“Ο«ΜαΒΎ“Μ ±ΦδΝΣœΒΘ©ΘΚ
’–ΤΗΘΚΡ‘”Αœώ ΐΨί¥ΠάμΙΛ≥Χ ΠΘ®÷Ί«λ&ΡœΨ©Θ©